
物種形成與生殖隔離機制:從障壁連乘到分子不相容
用族群遺傳學數學與分子動力學,拆解基因流如何被阻斷
從「停止交配」到「無法交配」:生殖隔離的本質
兩個族群什麼時候算是分成了兩個物種?直覺上我們會說「牠們不再交配」,但這只是表象。生殖隔離(reproductive isolation)真正的內涵是:兩個基因庫之間的基因流(gene flow)被某種機制阻斷,使得各自獨立累積差異。物種形成(speciation)的核心問題,因此不是「外觀差多少」,而是「能阻斷多少基因交換」。本篇從隔離機制的分子基礎、量化的隔離強度,以及推動分化的族群遺傳學數學出發,把這個過程拆解到可以計算的層次。
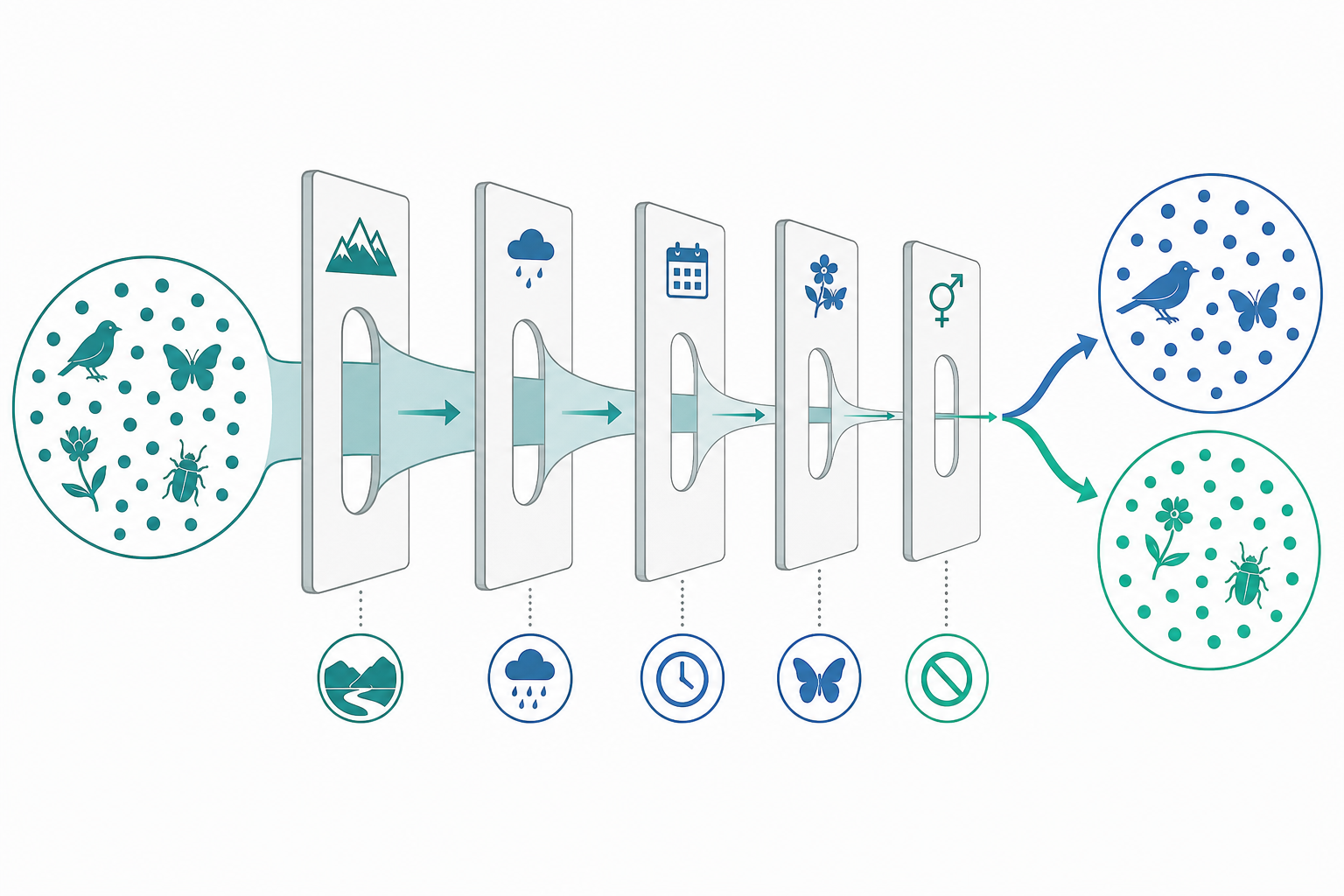
隔離機制的分類與量化
生殖隔離機制依作用時點分為合子前(prezygotic)與合子後(postzygotic)兩大類。合子前包括生態隔離、時間隔離、行為隔離、機械隔離與配子隔離;合子後包括雜種不活(hybrid inviability)、雜種不育(hybrid sterility)與雜種崩潰(hybrid breakdown)。
關鍵的觀念是:總隔離不是單一機制決定,而是序列性連乘。若各障壁讓基因流通過的比例為 $p_i$,則總通過率為 $\prod_i p_i$,總隔離強度為
$$RI_{\text{total}} = 1 - \prod_{i=1}^{n}(1 - RI_i)$$
其中 $RI_i$ 是第 $i$ 個障壁的個別隔離強度。Coyne 與 Orr 對果蠅與其他類群的經典分析指出,合子前障壁因為作用在前端,對阻斷基因流的「邊際貢獻」往往遠大於後端的合子後障壁——這也是為什麼強化作用(reinforcement)會優先增強交配前的辨識能力。
舉一個定量小範例。假設兩個同域族群之間有三道障壁:時間隔離 $RI_1 = 0.6$、行為(求偶辨識)隔離 $RI_2 = 0.8$、雜種不育 $RI_3 = 0.9$。則通過各障壁的比例分別為 $0.4$、$0.2$、$0.1$,總通過率 $= 0.4 \times 0.2 \times 0.1 = 0.008$,故
$$RI_{\text{total}} = 1 - 0.008 = 0.992.$$
注意若把行為隔離移除,總通過率躍升為 $0.4 \times 1 \times 0.1 = 0.04$,隔離強度掉到 $0.96$——前端障壁一旦失守,後端再強也補不回等量的基因流阻斷,這正是序列連乘的非線性後果。
合子後不相容的分子基礎:DM 模型
雜種為什麼會不育或不活?若某個位點的「壞」等位基因在親本背景中就有害,天擇早該把它清除,它根本傳不到雜種。Bateson–Dobzhansky–Muller(DM)不相容模型解決了這個悖論:分化發生在「不同位點」。族群 A 在位點 1 演化出新等位基因 $a$,族群 B 在位點 2 演化出新等位基因 $b$;$a$ 在 A 的遺傳背景中無害、$b$ 在 B 的背景中無害,但 $a$ 與 $b$ 從未在同一基因組中共存並接受檢驗。雜種第一次讓它們相遇,負向上位(epistasis)才暴露為不相容。
DM 模型有一個強力的數學預測:不相容是成對交互作用累積的結果,配對數隨分化位點數 $K$ 約以 $\binom{K}{2} \sim K^2/2$ 成長。因此雜種不相容應隨分化時間加速(snowball)累積,而非線性。Drosophila(如 D. melanogaster 的 Hmr、Lhr)與相關研究中陸續鑑定出的「物種形成基因」,許多涉及異染色質、轉錄調控與減數分裂衝突,符合此框架。
Hardy–Weinberg 基線與分化的偏離
要量化「分化」,必先有「沒有分化時應該長怎樣」的虛無模型。Hardy–Weinberg 平衡(HWE)給出隨機交配、無選汰、無突變、無遷移、無限大族群下的期望基因型頻率:
$$p^2 + 2pq + q^2 = 1, \quad p + q = 1.$$
物種形成的所有驅力——選汰、漂變、有限族群、非隨機交配——都是讓真實族群偏離 HWE 的力量。當兩個亞群因分化而各自走向不同等位基因頻率,整體相對於隨機交配會出現雜合子缺失(Wahlund 效應),可用固定指數量化:
$$F_{ST} = \frac{H_T - H_S}{H_T},$$
其中 $H_T$ 為合併族群期望雜合度、$H_S$ 為亞群平均期望雜合度。$F_{ST}$ 越高代表分化越深。全基因組掃描中常見的「分化島嶼」(islands of divergence)——即少數 $F_{ST}$ 異常高的區段,常對應到承載生殖隔離基因或受到強選汰的區域,是同域物種形成研究的核心證據。
驅動分化的數學:漂變、選汰與遷移的拉鋸
中性等位基因因遺傳漂變而固定的機率等於其當前頻率 $p$;新突變($p = 1/2N$,二倍體)固定機率為 $1/2N$,固定所需的平均時間約為 $4N_e$ 世代。族群越小,漂變越快,分化越容易啟動——這解釋了奠基者效應與周緣同域(peripatric)物種形成的威力。
但分化能否維持,取決於它與基因流的拉鋸。一個經典且關鍵的閾值是 $N_e m$(每世代有效遷移個體數):
$$N_e m \gtrsim 1 \implies \text{基因流足以抵抗漂變,分化難以維持。}$$
換言之,只要每世代平均有「一個以上」的有效遷移者,就足以把兩個亞群的中性差異抹平。要在基因流存在下仍完成物種形成(同域或近域),就必須有選汰強到足以克服遷移——這把我們帶向選汰係數 $s$ 與遷移率 $m$ 的競爭:唯有 $s > m$ 的位點,其分化才能在面對持續基因流時被選汰維持,這也是分化島嶼為何集中在強選汰區域的理論根據。
配子層次的隔離:一個生化視角
配子隔離常被當成黑箱,但它有扎實的分子動力學基礎。海膽 bindin、鮑魚 lysin 等配子辨識蛋白與卵子受體的結合具高度物種專一性,且這些蛋白常顯示正向選汰($d_N/d_S > 1$)的特徵。若把受精視為配子表面分子的結合反應,其有效結合速率對配子濃度的依賴,在形式上與酵素動力學同構——以 Michaelis–Menten 式
$$v = \frac{V_{\max}[S]}{K_M + [S]}$$
類比,異種配子間升高的「等效 $K_M$」(親和力下降)即對應到較低的受精效率。當辨識蛋白共演化使異種 $K_M$ 大幅升高,配子隔離強度便隨之增強。這提供了一條把「行為/生化辨識」嵌入前述 $RI_i$ 連乘框架的量化路徑。
深入探討(研究所視角)
在研究所層次,物種形成研究已從「描述障壁」轉向「在基因組與分子結構解析度上機制化隔離」。以下是主體未涵蓋的前沿面向。
結構生物學與共演化的軍備競賽。 配子辨識蛋白的正向選汰不只是統計現象。鮑魚 lysin 的晶體結構顯示其以單體—二聚體轉換驅動卵膜(VERL)溶解,而 VERL 含多個串聯重複序列,lysin 與 VERL 之間呈現界面共演化:一方關鍵殘基的取代會在介面上對另一方施加選汰壓。以結構為錨,研究者能把 $d_N/d_S$ 的訊號定位到實際的結合界面殘基,並用突變—結合自由能變化 $\Delta\Delta G$ 量化每個取代對親和力的貢獻——這把「物種專一性」從演化統計拉到了熱力學可測量。性衝突(sexual conflict)理論進一步預測,當受精動力學對雌性有成本時,雌性受體會演化以「降低」結合效率,形成持續的拮抗共演化,恰可解釋這些蛋白異常快速的演化速率。
單細胞與體學重寫雜種不相容。 傳統雜種不育研究止於「雄性不育」這類表型,單細胞 RNA 定序則能解析不育發生在減數分裂的哪一個細胞狀態。在果蠅與小鼠雜種睪丸的單細胞圖譜中,可觀察到特定減數分裂階段的轉錄程序崩潰、以及減數分裂性染色體去活化(MSCI)失效導致 X 連鎖基因異常表達——這為「大 X 效應」與 Haldane 法則(異配性別的雜種優先受害)提供了機制層面的解釋,而非僅是統計規律。ATAC-seq 等染色質可及性體學則揭示,許多不相容根源於順式調控元件與反式作用因子(如 KRAB-zinc finger 蛋白與其標靶的轉位子)之間的快速共演化失配。
減數分裂驅動與基因組衝突作為物種形成引擎。 近年一個重要轉向,是把生殖隔離基因的快速演化歸因於內部基因組衝突而非外部生態適應。自私遺傳元件——分離扭曲子(segregation distorters)、減數分裂驅動的著絲點、轉位子——與壓制它們的宿主因子之間的紅皇后式軍備競賽,會驅動異染色質蛋白與小 RNA(piRNA)路徑的快速分化。當兩物種各自演化出不同的「驅動—壓制」配對,雜種中的錯配即表現為不育或分離異常,這正是 DM 不相容的一個具體分子實現。
系統生物學的網路觀點。 DM 不相容本質上是負向上位,而上位的結構由基因調控網路(GRN)決定。以網路模型模擬,可顯示不相容的 snowball 累積速率取決於網路的連結度與模組性:高度互連的網路產生更陡的不相容曲線。將群體遺傳模擬(如擴散近似下的等位基因頻率動態)與機制性 GRN 耦合,正在成為預測「哪些位點、在多少世代後、以多快速度」貢獻生殖隔離的整合框架——讓物種形成從一個歷史敘事,逐步變成可被參數化、可被檢驗的定量科學。