
一個細胞變兩個:有絲分裂與減數分裂的生命密碼
從受精卵到 37 兆個你,再到孩子為何獨一無二——看懂生長、修復與遺傳變異的根源。
一個細胞變兩個,這件事為什麼這麼重要?
你身上大約有 37 兆個細胞,但你最初只是一個受精卵——一顆肉眼幾乎看不見的小球。從那一顆細胞長成現在的你,靠的就是「細胞分裂」:一個細胞變成兩個、兩個變成四個,不斷複製下去。
不只是長大。你今天劃傷手指,幾天後傷口癒合;你的腸壁細胞每幾天就換一批;你的紅血球壽命大約三個月就得補充。這些「修復」與「更新」全都靠細胞分裂。更神奇的是,當生命要把基因傳給下一代時,用的是另一種特別的分裂——它不只複製,還會「洗牌」,創造出獨一無二的後代。
這篇文章要帶你看懂兩種分裂:負責「長大與修復」的有絲分裂(mitosis),以及負責「生殖與變異」的減數分裂(meiosis)。先把核心觀念講清楚,再一路深入到大學普通生物學的程度。
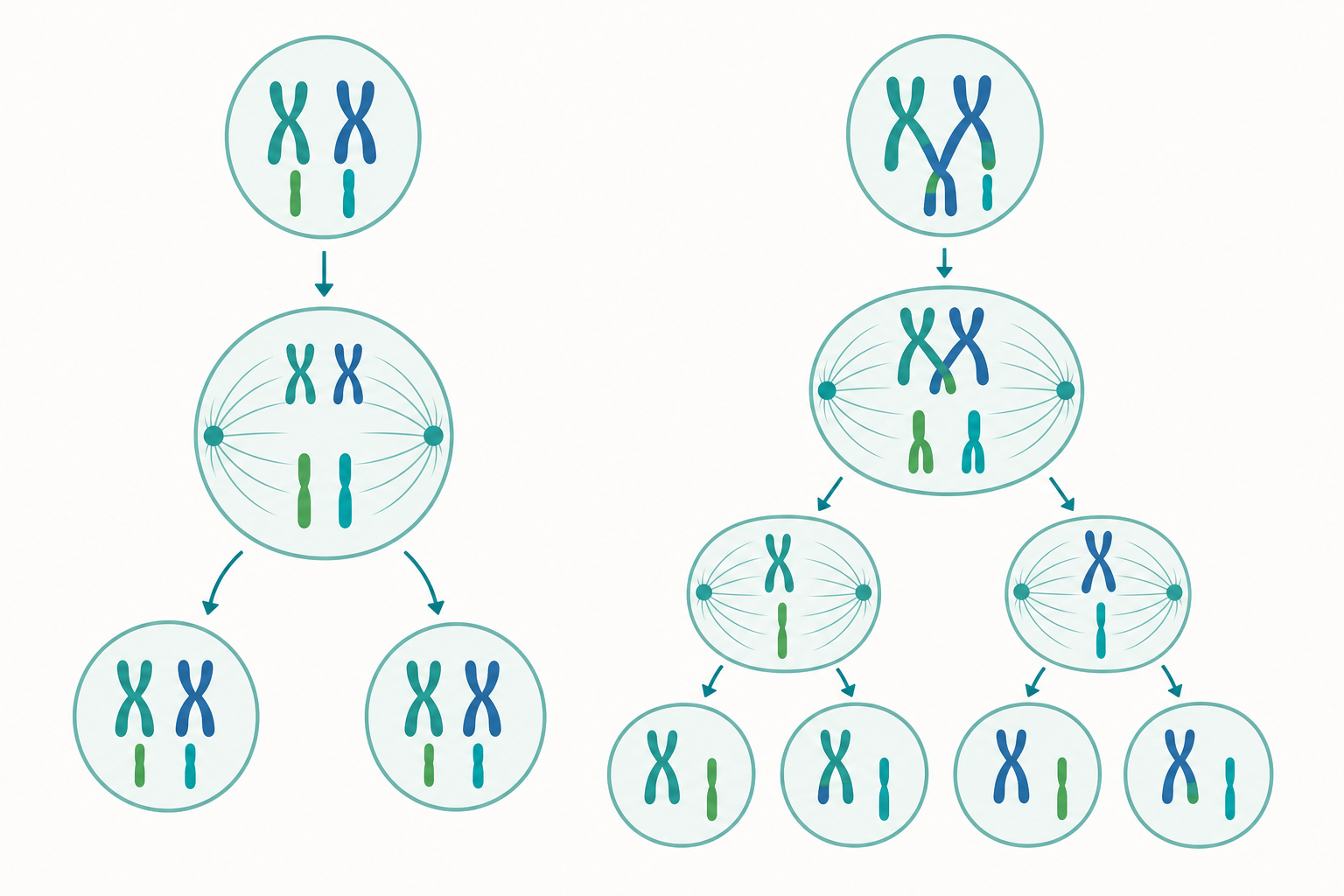
有絲分裂:印出兩份一模一樣的複本
想像你有一本重要的筆記本,要做一份完全相同的副本。你會怎麼做?先「影印」每一頁,再把兩份分開裝訂。細胞做的事幾乎一樣。
細胞分裂前,DNA 必須先完整複製一份,這發生在所謂的 S 期(合成期)。複製後,每一條染色體都變成「兩條姊妹染色分體」連在一起,像一個 X 形。整個細胞週期可以簡單記成:
$$G_1 \rightarrow S \rightarrow G_2 \rightarrow M$$
其中 $G_1$、$G_2$ 是生長與準備期,$S$ 是 DNA 複製期,$M$ 才是真正的「分裂」期。
有絲分裂(M 期)通常分成四個階段,可以這樣對照記:
- 前期(Prophase):染色質凝聚成看得見的染色體,核膜開始瓦解。
- 中期(Metaphase):所有染色體整齊排到細胞正中央的「赤道板」。
- 後期(Anaphase):姊妹染色分體被紡錘絲拉開,往兩極移動。
- 末期(Telophase):兩端各自重新形成核膜,接著細胞質分裂成兩個細胞。
結果是:一個母細胞變成兩個基因完全相同的子細胞,染色體數目不變。如果母細胞是人類體細胞(46 條),兩個子細胞也都是 46 條。這就是為什麼你皮膚再生出來的還是皮膚、肝臟修復出來的還是肝臟——複本忠實得一字不差。
減數分裂:為什麼孩子不會跟父母一模一樣?
如果生殖細胞也用有絲分裂,會出大問題。假設精子有 46 條染色體、卵子也有 46 條,受精後豈不是變成 92 條?下一代再翻倍成 184 條……幾代下來就失控了。
大自然的解法是減數分裂:在製造精子和卵子(合稱配子)時,染色體數目要先「減半」。人類配子只有 23 條,精卵結合後才回到 46 條。
減數分裂最關鍵的是它有兩次連續分裂(減數第一分裂與第二分裂),但 DNA 只複製一次。我們用一個簡單對照看清差異:
| 比較項目 | 有絲分裂 | 減數分裂 |
|---|---|---|
| 分裂次數 | 1 次 | 2 次 |
| 產生子細胞數 | 2 個 | 4 個 |
| 染色體數變化 | 不變(2n→2n) | 減半(2n→n) |
| 子細胞是否相同 | 完全相同 | 各不相同 |
| 功能 | 生長、修復 | 製造配子、生殖 |
更妙的是減數分裂會製造「變異」,靠兩個機制:
第一是聯會與互換(crossing over):在減數第一分裂前期,來自父方與母方的同源染色體會配對靠攏,彼此交換一段 DNA,像兩條繩子交換了一截。
第二是獨立分配(independent assortment):每一對同源染色體要往哪一極走,是隨機的。人有 23 對染色體,光是這個隨機組合就有 $2^{23} \approx 838$ 萬種可能。再加上互換,幾乎不可能有兩個配子完全相同——這就是為什麼除了同卵雙胞胎,世界上沒有兩個人基因一樣。
用孟德爾的豌豆,看懂變異怎麼來
減數分裂的「獨立分配」正是孟德爾遺傳定律的細胞基礎。我們用經典的單性狀雜交算一次。
假設高莖($T$)對矮莖($t$)為顯性。讓純種高莖($TT$)和純種矮莖($tt$)雜交:
- 第一代($F_1$):每個後代都拿到一個 $T$、一個 $t$,基因型全是 $Tt$,外表全為高莖。
- 第二代($F_2$):讓 $Tt \times Tt$ 自交。因為減數分裂把 $T$ 和 $t$ 分到不同配子,每個配子帶 $T$ 或 $t$ 的機率各半。
把配子組合畫成棋盤:
$$ \begin{array}{c|cc} & T & t \\ \hline T & TT & Tt \\ t & Tt & tt \\ \end{array} $$
結果基因型比例是 $1\,TT : 2\,Tt : 1\,tt$;因為 $T$ 是顯性,外表(表現型)比例就是高莖 : 矮莖 = 3 : 1。
請注意這個 3:1 不是巧合,而是直接來自減數分裂「等位基因分離、配子隨機結合」的結果。孟德爾在還不知道染色體與 DNA 的年代,就從豌豆數據推出了這個規律——後來細胞學發現的減數分裂,恰好解釋了他的定律為什麼成立。
當分裂失控:把概念連到健康
正常細胞知道「什麼時候該分裂、什麼時候該停」,這由一套嚴密的檢查機制把關。一旦控制系統出錯,細胞可能不停分裂、不肯停下,這就是癌症最核心的特徵之一。理解細胞週期,正是理解許多疾病與治療(例如化療藥物常針對快速分裂的細胞)的起點。反過來,若減數分裂時染色體沒分好(不分離,nondisjunction),配子就會多一條或少一條染色體,可能造成如唐氏症(多一條第 21 號染色體)等狀況。
深入探討(研究所視角)
進入分子層次,細胞週期的核心引擎是週期素(cyclin)與週期素依賴型激酶(CDK)的週期性配對。CDK 本身是激酶,但需與特定 cyclin 結合才有活性;不同 cyclin 在週期不同階段累積與被降解,使 CDK 活性呈波形震盪,驅動細胞依序通過各期。Leland Hartwell、Paul Nurse 與 Tim Hunt 因闡明此調控機制獲 2001 年諾貝爾生理醫學獎。其中 $G_1/S$ 與 $G_2/M$ 等檢查點(checkpoints)負責驗證 DNA 完整性與複製是否就緒;DNA 損傷會啟動 ATM/ATR–CHK 訊息級聯,活化 p53,誘導 p21 抑制 CDK,使週期停滯以爭取修復時間。p53 因此被稱為「基因組守衛者」,其失活是多數人類腫瘤的共同特徵。
紡錘體組裝檢查點(SAC)是另一關鍵:在後期啟動前,SAC 監測每個著絲點(kinetochore)是否都被來自兩極的微管正確且雙向附著(amphitelic attachment)。只要有一個著絲點尚未張力到位,SAC 便抑制後期促進複合體(APC/C),阻止 securin 被降解、separase 被活化,從而延後姊妹染色分體分離。這套機制的失靈正是染色體不穩定性(CIN)與非整倍體(aneuploidy)的來源,與腫瘤惡性度及減數不分離皆有關。
減數分裂的分子細節更為精巧。互換並非隨機斷裂,而是由 SPO11 蛋白主動製造 DNA 雙股斷裂(DSB)所引發,再經同源重組(以同源染色體而非姊妹染色分體為模板)修復,形成 Holliday 接合並解離為互換產物。聯會複合體(synaptonemal complex)穩定同源配對,而交叉干擾(crossover interference)確保每對同源染色體至少有一個交叉(obligate crossover),這是正確分離的物理前提。減數第一分裂中,姊妹著絲點被 shugoshin–PP2A 保護而「協同附著」(朝同一極),與有絲分裂的雙向附著截然不同——這個差異正是「同源染色體先分、姊妹分體後分」兩段式分裂的分子基礎。
研究前沿上,單細胞定序讓人能對單一配子或胚胎重建個體化的重組與不分離圖譜,量化人類減數分裂的錯誤率與年齡效應(如高齡卵母細胞 cohesin 流失導致不分離上升);CRISPR–Cas9 則使研究者能精準擾動 SAC、cohesin 或檢查點基因以驗證因果。將上述調控網絡放入系統生物學框架,可用常微分方程描述 cyclin–CDK 的雙穩態與遲滯(hysteresis),把「細胞為何只在跨越閾值後才不可逆地進入分裂」這件事,從定性敘述提升為可預測的動力學模型——這正是當代細胞週期研究從分子清單走向定量機制的方向。