
細胞裡的化學魔法師:酵素如何用一條捷徑掌控生命
從活化能、鎖鑰契合到代謝路徑,看酵素如何讓上千個反應井然有序地發生
細胞裡的「化學魔法師」:先認識酵素
想像你站在一座高高的山丘前,山的另一邊放著你想要的東西。如果直接翻山,你得花費大把力氣爬到山頂才下得去;但如果有人替你在山中鑿了一條隧道,你幾乎不費吹灰之力就能穿過去。在細胞的世界裡,這條「隧道」就是酵素(enzyme)所做的事。
細胞裡每一秒都在發生成千上萬個化學反應:把食物分解成能量、把胺基酸組裝成蛋白質、複製 DNA、清除廢物。這些反應若放在試管裡,多數都慢得驚人,有些甚至需要數百年才能完成一次。但在細胞裡,它們在毫秒之間就完成了。讓速度暴增的關鍵,正是酵素——一種多半由蛋白質構成的生物催化劑(biocatalyst)。
酵素最迷人的地方在於:它讓反應「變快」,自己卻「不被消耗」。一個酵素分子可以一遍又一遍地工作,像一位不知疲倦的工匠,重複地把原料變成產品。
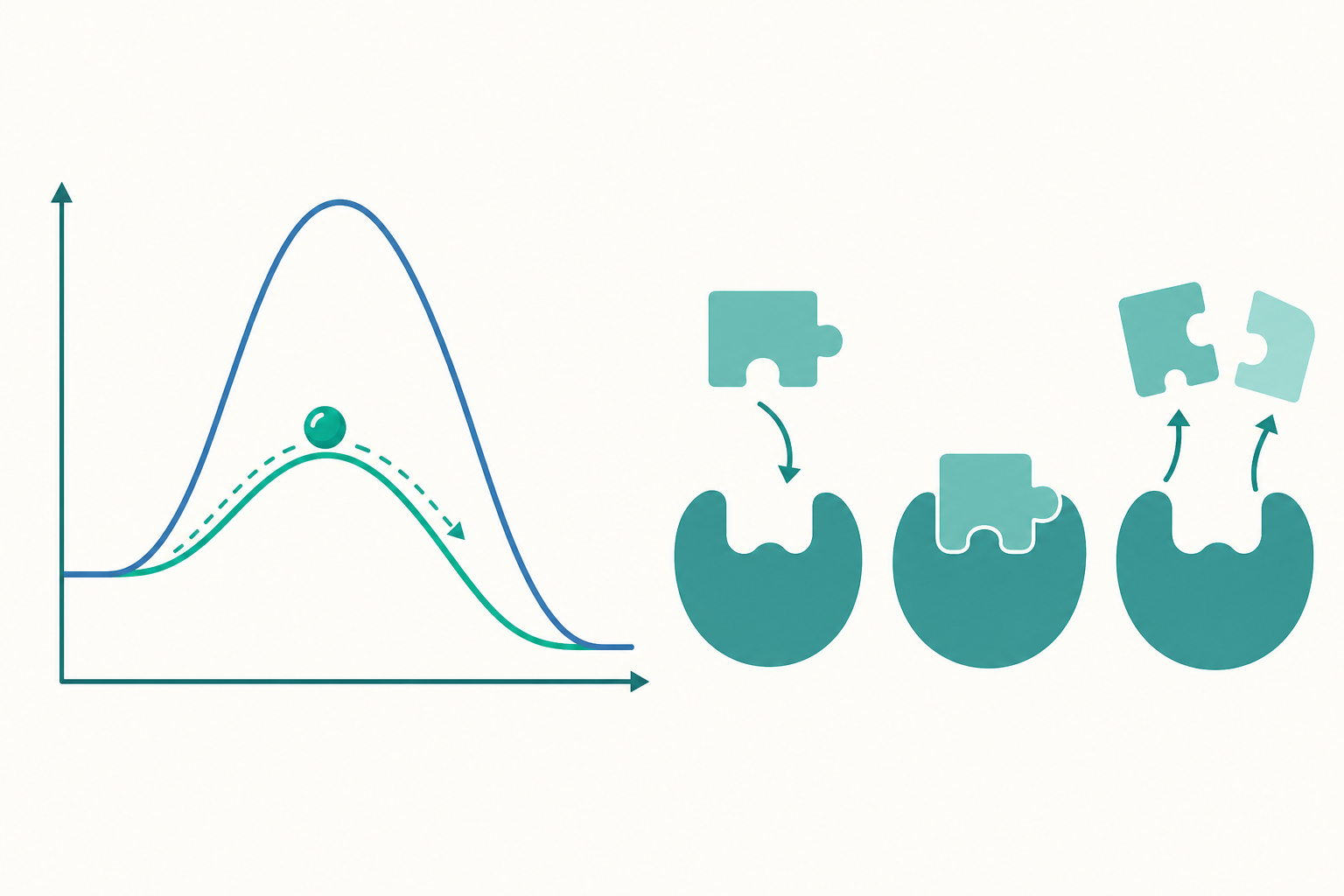
活化能:那座非翻不可的山
要理解酵素為什麼厲害,得先認識一個概念:活化能(activation energy, $E_a$)。
任何化學反應的進行,反應物都必須先越過一個「能量門檻」,才能轉變成產物。這個門檻就是活化能,也就是前面比喻裡那座山的高度。山愈高,能翻過去的分子愈少,反應就愈慢。
酵素的本領,就是降低活化能。它並不會改變反應最終釋放或吸收的總能量(也就是山兩側的高度差不變),而是替反應「鑿了一條更低的路徑」。我們可以用一個直覺的關係來理解反應速率與活化能:
$$k = A \, e^{-E_a / RT}$$
這是著名的阿瑞尼士方程式(Arrhenius equation)。其中 $k$ 是反應速率常數,$E_a$ 是活化能,$R$ 是氣體常數,$T$ 是絕對溫度。從式子可以看出:當酵素把 $E_a$ 變小,指數項 $e^{-E_a/RT}$ 就變大,反應速率 $k$ 隨之大幅提升。哪怕只把活化能降低一點點,速率也可能增加成千上萬倍。
值得強調的是:酵素只是「鋪了一條捷徑」,它無法讓本來不會發生的反應變成會發生。熱力學上不利的反應,酵素改變不了結局,它只能改變抵達結局的速度。
鎖與鑰匙:專一性從哪裡來
酵素另一個關鍵特性是專一性(specificity):一種酵素通常只催化一種或一類反應。負責分解澱粉的澱粉酶不會去碰脂肪,分解蛋白質的胃蛋白酶也不會去動醣類。
這份專一性來自酵素表面一個特殊的凹槽,叫做活性位(active site)。反應物(稱為受質,substrate)會嵌進這個凹槽,就像鑰匙插進鎖孔。早期科學家用「鎖與鑰匙模型」來形容這種精準契合。
不過後來的研究發現,事情更靈活一些:當受質靠近時,酵素的形狀會稍微調整,把受質「抱」得更緊。這稱為誘導契合模型(induced-fit model)。可以把它想成握手——你的手不是僵硬的模子,而是會順著對方的手調整握力。
一次完整的酵素作用可以用這條流程表示:
$$E + S \;\rightleftharpoons\; ES \;\longrightarrow\; E + P$$
其中 $E$ 是酵素,$S$ 是受質,$ES$ 是兩者結合的中間複合物,$P$ 是產物。注意最右邊:酵素 $E$ 完好地回來了,準備迎接下一個受質。
影響酵素活性的三個旋鈕
酵素的工作效率不是固定的,至少有三個因素像旋鈕一樣調節它:
-
溫度:溫度升高,分子碰撞變頻繁,反應加快——但只到某個點為止。超過最適溫度,蛋白質結構會被破壞(稱為變性,denaturation),活性急速下降。人體酵素的最適溫度大約在攝氏 37 度,這也正是體溫的數值,並非巧合。
-
酸鹼值(pH):每種酵素都有偏好的酸鹼環境。胃蛋白酶在強酸的胃裡(pH 約 2)最活躍;而小腸裡的胰蛋白酶則偏好微鹼環境(pH 約 8)。離開最適 pH,活性同樣會下滑。
-
受質濃度:受質愈多,反應一開始愈快;但當所有酵素分子都「忙碌中」時,速率會達到上限,不再增加。這個飽和現象是酵素動力學的核心,後面研究所段落會更深入。
把酵素串起來:代謝路徑
單一酵素已經很神奇,但細胞真正的高明之處,是把許多酵素串接成生產線,這就是代謝路徑(metabolic pathway)。
以我們吃下的葡萄糖為例。葡萄糖不會「砰」地一聲直接變成能量,而是經過一連串小步驟,每一步由不同的酵素催化,像工廠流水線一樣,前一站的產物是下一站的原料。我們熟悉的細胞呼吸總反應式是:
$$C_6H_{12}O_6 + 6\,O_2 \;\longrightarrow\; 6\,CO_2 + 6\,H_2O + \text{能量(ATP)}$$
這條簡潔的式子背後,其實藏著糖解作用、檸檬酸循環、電子傳遞鏈等數十個由酵素催化的步驟。
為什麼要分這麼多步?因為這樣細胞才能精準控制。在生產線的某一站放慢或加速,整條路徑的產出就會跟著改變。一個常見的調控方式叫做回饋抑制(feedback inhibition):當終產物累積太多時,它會回頭抑制路徑前段的某個酵素,就像水庫滿了就關上閘門,避免資源浪費。這正是「酵素如何調控細胞中成千上萬反應」的縮影——不是靠蠻力,而是靠精巧的開關與節流。
為什麼這一切重要
酵素不只是課本上的名詞。發酵釀酒、用洗衣酵素去除汙漬、乳糖不耐症(缺乏分解乳糖的乳糖酶)、許多藥物其實是「酵素抑制劑」——這些日常現象的核心都是酵素。理解酵素如何降低活化能、如何被調控,就等於拿到了一把理解生命運作與生物科技的鑰匙。
深入探討(研究所視角)
入門段落用「速率有上限」一筆帶過了酵素的飽和行為,這裡補上其定量骨架。1913 年提出的 Michaelis–Menten 動力學將反應初速率 $v$ 描述為:
$$v = \frac{V_{\max}[S]}{K_m + [S]}$$
其中 $V_{\max}$ 是酵素被受質飽和時的最大速率,$K_m$(米氏常數)在數值上等於 $v = V_{\max}/2$ 時的受質濃度。$K_m$ 常被當作酵素對受質「親和力」的指標:$K_m$ 愈小,表示低濃度受質就能讓酵素半飽和,親和力愈高。此模型建立在穩態假設(steady-state assumption,Briggs–Haldane)之上,即 $ES$ 複合物的濃度在反應初期近似恆定。實驗上常用 Lineweaver–Burk 雙倒數作圖將其線性化,以便從截距與斜率估計 $V_{\max}$ 與 $K_m$,但此法會放大低濃度區的誤差,現代多改用非線性回歸直接擬合。
抑制劑的分類也比入門所述精細:競爭性抑制劑與受質爭奪活性位,使表觀 $K_m$ 上升而 $V_{\max}$ 不變;非競爭性抑制劑結合在活性位以外的位點,降低 $V_{\max}$ 而不改變 $K_m$。許多臨床藥物正是依此設計,例如他汀類藥物即為 HMG-CoA 還原酶的競爭性抑制劑。
更進階的是異位調控(allosteric regulation)。許多代謝關鍵酵素具有多個次單元與調節位點,其速率曲線呈 S 型(sigmoidal)而非雙曲線,可用 Hill 方程式描述協同效應。回饋抑制往往透過異位機制實現,使酵素在受質濃度的狹窄區間內如「開關」般靈敏切換——這是線性 Michaelis–Menten 模型無法捕捉的非線性行為。
在催化機制層面,酵素降低活化能的物理本質是穩定反應的過渡態(transition state):活性位與過渡態的結合能比與受質的結合能更強,差額即用以抵消活化能障壁。這個洞見啟發了過渡態類似物作為強效抑制劑,以及人工設計的催化抗體(abzymes)。
連結到更前沿的研究:系統生物學運用通量平衡分析(flux balance analysis)對全基因組規模的代謝網路建模,預測在不同營養或基因敲除條件下的代謝通量重分配;單細胞代謝體學與單細胞定序則揭露同一群細胞間酵素表現的異質性;而 CRISPR 基因編輯讓研究者能精準敲除或調控特定酵素基因,直接驗證其在代謝網路中的角色。酵素,從一條「化學捷徑」的比喻,一路通向定量模型、藥物設計與系統層級的生命工程。