
從原子到行星:生物地球化學循環與全球變遷的量化視角
以駐留時間、酶動力學、氧化還原電位與化學計量比,解構碳氮磷循環如何驅動氣候系統
從原子到行星:生物地球化學循環的量化視角
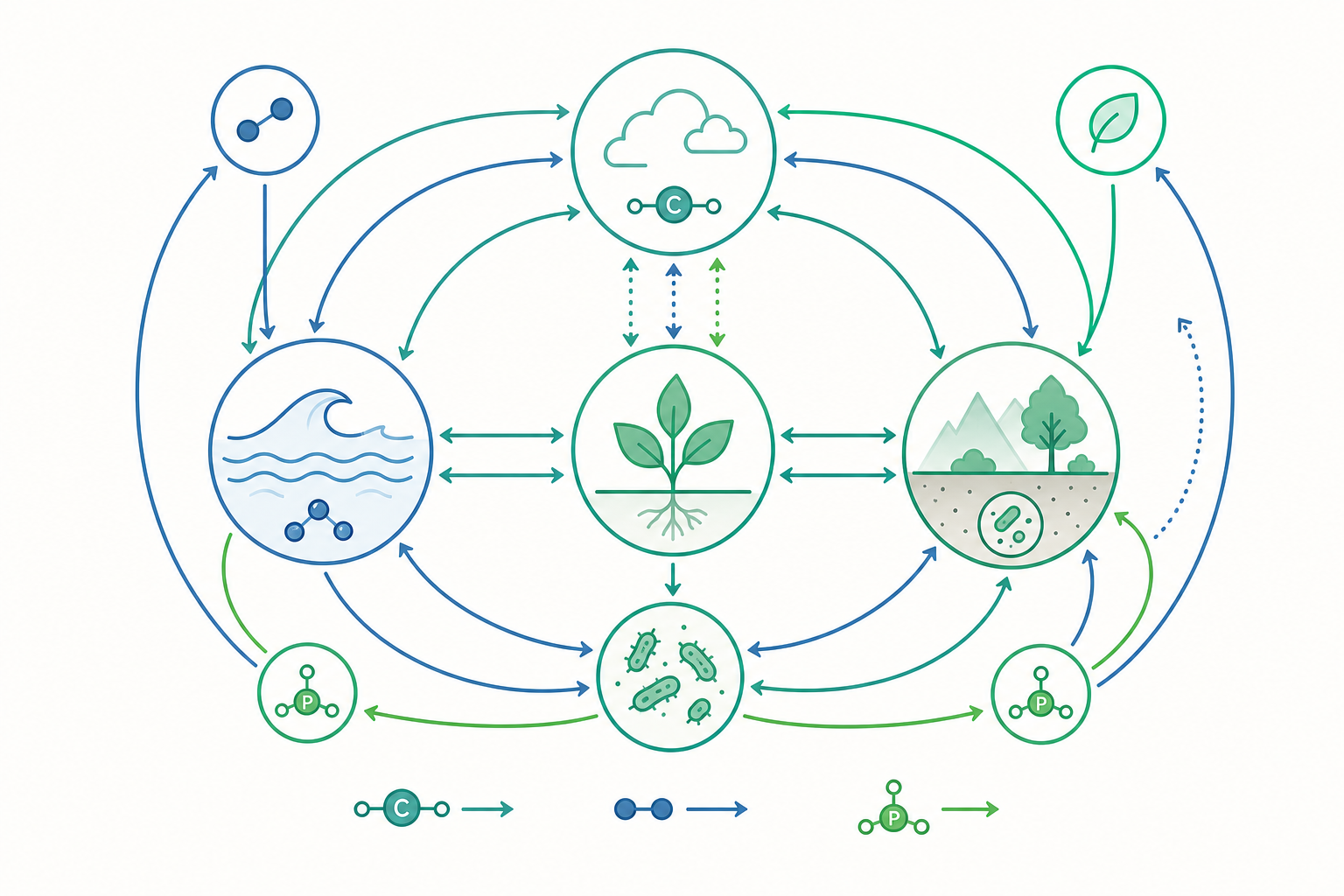
地球上的碳、氮、磷、硫等元素並非被生命「消耗」掉,而是在岩石圈、水圈、大氣圈與生物圈之間不停地被搬運、轉化與儲存。每一個原子在某個「庫(reservoir)」裡停留一段平均時間,再隨著某個「通量(flux)」移動到下一個庫。把這些庫與通量畫成一張收支表,就是生物地球化學循環的本質——而當人類在百年尺度上改變了通量,全球變遷便隨之發生。
要量化一個元素在某庫中的駐留時間(residence time, $\tau$),可用一個極簡卻深刻的關係式:
$$\tau = \frac{M}{F_{\text{out}}}$$
其中 $M$ 為庫存量(mass),$F_{\text{out}}$ 為總流出通量。以大氣中的 CO$_2$ 為例,工業化前大氣碳庫約 $600\ \text{Pg C}$,年總流出(光合作用 + 海洋吸收)約 $200\ \text{Pg C·yr}^{-1}$,故 $\tau \approx 3$ 年——這指的是「單一分子」的快速週轉。但一旦把海洋深層與岩石風化納入,「過量」CO$_2$ 真正被移除的時間尺度卻拉長到數百至數千年,這個快慢時間尺度的不一致,正是氣候惰性的根源。
碳循環:從 RuBisCO 的動力學瓶頸談起
陸地碳匯的速率,最終受制於一個分子層級的瓶頸:核酮糖-1,5-雙磷酸羧化酶/加氧酶(RuBisCO)。它同時是地球上最豐富的蛋白質,也是效率最差的主要酶之一,$k_{\text{cat}}$ 僅約每秒 3–10 次。它的固碳速率可用 Michaelis–Menten 方程描述:
$$v = \frac{V_{\max}\,[\text{CO}_2]}{K_m + [\text{CO}_2]}$$
C3 植物 RuBisCO 對 CO$_2$ 的 $K_m$ 約 $10\text{–}15\ \mu\text{M}$,與當前大氣對應的葉肉 CO$_2$ 濃度相近,意味著酶尚未飽和。因此大氣 CO$_2$ 上升時,$v$ 仍會增加——這就是「CO$_2$ 施肥效應(CO$_2$ fertilization)」的分子基礎。然而 RuBisCO 的致命缺陷是同時催化加氧反應(光呼吸),而溫度升高會使其對 O$_2$ 的相對親和力上升,削弱固碳。C4 植物與 CAM 植物則演化出 CO$_2$ 濃縮機制,在 RuBisCO 周圍局部提高 $[\text{CO}_2]$,把工作點推向 $V_{\max}$,這是對動力學瓶頸的生理層級突圍。
定量小範例:設某 C3 葉片 $V_{\max}=100\ \mu\text{mol·m}^{-2}\text{·s}^{-1}$,$K_m=12\ \mu\text{M}$。若葉肉 $[\text{CO}_2]$ 由 $8\ \mu\text{M}$(工業化前)升至 $14\ \mu\text{M}$:
$$v_1=\frac{100\times 8}{12+8}=40,\qquad v_2=\frac{100\times 14}{12+14}\approx 53.8$$
固碳速率提升約 $(53.8-40)/40 \approx 35\%$。但注意這只是「潛在」增幅,實際受氮、磷、水分共同限制(Liebig 最小因子律),這正是地球系統模型對碳匯預測高度不確定的原因。
氮循環:人類已改寫的全球氮平衡
氮循環是人類擾動最劇烈的循環。工業固氮(Haber–Bosch 法)每年將約 $120\ \text{Tg N}$ 由 N$_2$ 轉為活性氮,已超過所有陸地天然固氮的總和。微生物驅動的各步驟——固氮、硝化、反硝化、厭氧氨氧化(anammox)——本質上都是氧化還原反應,可用 Nernst 方程理解其能量驅動力:
$$E = E^{\circ} - \frac{RT}{nF}\ln Q$$
不同氮形態(NH$_4^+$/NO$_2^-$/NO$_3^-$/N$_2$)對應不同的標準還原電位,微生物會優先利用能釋放最多自由能的電子受體序列(O$_2$ > NO$_3^-$ > Mn$^{4+}$ > Fe$^{3+}$ > SO$_4^{2-}$)。自由能與電位的關係為:
$$\Delta G = -nF\,\Delta E$$
當土壤或水體缺氧,反硝化作用把 NO$_3^-$ 還原為 N$_2$ 與 N$_2$O。後者是溫室效應為 CO$_2$ 約 273 倍的長壽溫室氣體,也是平流層臭氧的主要破壞者。過量施肥導致的活性氮逕流,最終在河口造成優養化與低氧區(hypoxic zone),構成從分子電位到行星尺度的因果鏈。
磷循環、化學計量比與全球變遷的耦合
與碳、氮不同,磷沒有顯著的氣相,主要循環受岩石風化與沉積控制,週轉極慢。海洋浮游生物的元素組成長期維持在著名的 Redfield 比 $C:N:P \approx 106:16:1$,這個近乎恆定的化學計量比,把三個循環緊密耦合:固定一定量的碳,就「鎖定」了相應比例的氮與磷需求。
因此磷常是陸地與海洋初級生產的終極限制因子。當全球暖化改變海洋分層與上湧,營養鹽供應比例偏離 Redfield 比,就會重塑浮游群落結構與生物碳泵(biological carbon pump)的效率——這是循環之間「牽一髮動全身」的系統性風險。
深入探討(研究所視角)
在分子與系統層級,當代地球化學的前沿正快速重寫教科書圖像。RuBisCO 工程是其中焦點:結構生物學透過 X 光晶體繞射與冷凍電鏡解析其大/小次單元的活性中心,揭示了 carbamylation(活化離胺酸與 CO$_2$ 形成胺甲酸酯,再由 Mg$^{2+}$ 穩定)的精確幾何。研究者試圖移植藍綠菌的羧酶體(carboxysome)或藻類的蛋白核(pyrenoid)這類 CO$_2$ 濃縮構造進入 C3 作物,並借助定向演化篩選更高 $k_{\text{cat}}$ 或更低加氧/羧化比($\Omega$ 因子)的變異株,目標是在分子層級鬆開全球碳匯的瓶頸。
單細胞與體學正解構過去被視為「黑盒」的微生物地球化學。透過 nanoSIMS(奈米級二次離子質譜)結合穩定同位素標定,可在單一細胞尺度直接觀測 $^{15}$N$_2$ 或 $^{13}$CO$_2$ 的同化速率,量化個別細胞對總通量的貢獻;而宏基因組(metagenomics)與宏轉錄組則揭示了 comammox(完全氨氧化於單一 Nitrospira 菌株內完成)的存在,徹底顛覆了硝化作用必由兩類菌分工的百年範式。anammox 菌的厭氧氨氧化體(anammoxosome)內以聯氨(hydrazine, N$_2$H$_4$)為中間產物的代謝,更被視為對「不可能化學」的生物學解。
系統生物學與地球系統模型(ESM)則嘗試把這些分子細節向上整合至行星尺度。挑戰在於跨越十餘個數量級的時空尺度耦合:酶動力學(毫秒、奈米)如何約束生態系淨交換(年、公里)。當代模型如 CMIP6 框架下的耦合碳—氣候模擬,已將氮、磷限制顯式納入(C–N–P coupling),因為忽略養分限制會系統性高估未來陸地碳匯。一個關鍵且尚未閉合的問題是「碳—氣候回饋」的正負號與強度:暖化加速土壤微生物呼吸(受 Arrhenius 溫度依賴性 $k \propto e^{-E_a/RT}$ 支配,常以 $Q_{10}$ 參數化),可能把陸地由碳匯翻轉為碳源。
近期最受關注的機制發現,包括北方凍土融解釋放古老有機碳、海洋「生物碳泵」中顆粒沉降與微生物再礦化的競爭,以及鐵作為高營養低葉綠素(HNLC)海域限制因子的角色——這些都把微觀的酶與電子傳遞鏈,與宏觀的冰期—間冰期 CO$_2$ 振盪連結起來。研究所層級的核心訓練,正是學會在這些尺度之間,以化學計量、熱力學與動力學為共通語言,建立可被同位素與通量觀測檢驗的定量假說。