
從一顆受精卵到精準身體:形態發生的數理邏輯
形態原梯度、反應—擴散與基因調控網絡如何把化學濃度轉譯成精確的身體模式
從一顆受精卵到精準身體:形態發生的數理邏輯
一顆受精卵如何「知道」哪裡該長出頭、哪裡該長出尾?答案不在某個神祕的設計藍圖,而在分子濃度的空間梯度、基因網絡的開關邏輯,以及細胞之間力學與化學的反覆對話。發育生物學的核心命題,正是把這種「看似有意志的自我組織」拆解成可量化的反應—擴散、訊號傳遞與機械力學過程。
在普通生物學裡,我們學到囊胚、原腸化、三胚層;但真正的發育生物學處理的是模式形成(pattern formation)的精確度問題:果蠅胚胎能在約三小時內、以單一細胞寬度的解析度劃分出體節邊界,其位置誤差小於 1%。要理解這種精度,必須引入濃度梯度、隨機性與基因調控網絡的數學語言。
形態原梯度與位置資訊
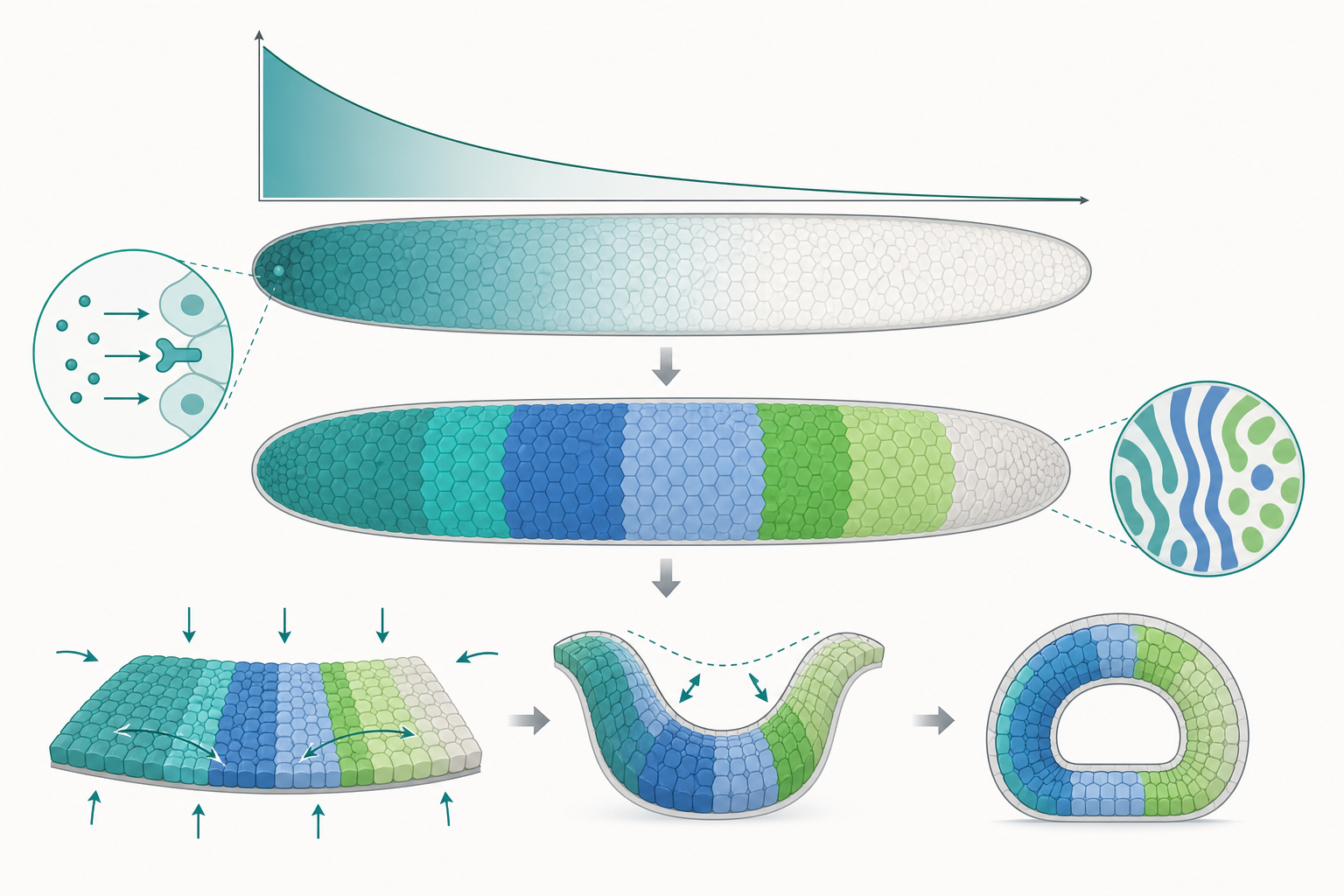
Lewis Wolpert 在 1969 年提出的位置資訊(positional information)假說主張:細胞依據其所處的形態原(morphogen)濃度,讀取自己在胚胎中的座標,進而決定命運。最經典的分子實證是果蠅母系效應基因 bicoid。Bicoid 蛋白由胚胎前端的 mRNA 翻譯後向後擴散,並持續降解,形成一條由前到後遞減的指數型梯度。
在最簡單的合成—擴散—降解(SDD)模型中,穩態濃度滿足:
$$ D \frac{\partial^2 C}{\partial x^2} - k\,C = 0 $$
其中 $D$ 為擴散係數、$k$ 為降解速率常數。解為指數衰減:
$$ C(x) = C_0\, e^{-x/\lambda}, \qquad \lambda = \sqrt{D/k} $$
特徵長度 $\lambda$ 決定梯度的「陡峭程度」。以 Bicoid 為例,實測 $\lambda$ 約 100 微米,恰好涵蓋胚胎前半部,使下游目標基因(如 hunchback)能在特定閾值處被切換開啟。這正是形態原把連續的化學濃度,轉譯成離散的基因表現域的方式。
定量小範例:閾值讀取的位置精度
假設 hunchback 在 Bicoid 濃度降到初始值 $C_0$ 的 $1/e$(約 37%)時被啟動。求啟動邊界的位置 $x^*$。
由 $C(x^*) = C_0 e^{-x^*/\lambda} = C_0/e$,可得 $e^{-x^*/\lambda} = e^{-1}$,故 $x^* = \lambda \approx 100\ \mu m$。
接著考慮精度:若梯度幅度有 10% 的胚胎間變異($\Delta C/C = 0.1$),對應的位置誤差為
$$ \Delta x = \lambda\,\frac{\Delta C}{C} = 100 \times 0.1 = 10\ \mu m $$
相當於約 2–3 個細胞寬度。實際胚胎卻能達到單細胞精度,這說明系統不能只靠單一梯度——它必須整合多個基因的交叉調控與時間平均,才能壓低雜訊。這個落差,正是 1990 年代以降發育定量研究的核心驅動力。
反應—擴散:圖靈的自組織機制
Alan Turing 在 1952 年提出,兩種擴散速率不同、彼此調控的化學物質(活化子 activator 與抑制子 inhibitor)即可從均勻初始狀態自發產生週期性空間圖案。其線性化系統可寫為:
$$ \frac{\partial u}{\partial t} = f(u,v) + D_u \nabla^2 u, \qquad \frac{\partial v}{\partial t} = g(u,v) + D_v \nabla^2 v $$
關鍵條件是抑制子擴散遠快於活化子($D_v \gg D_u$),即「局部自我活化、長程抑制」。當均勻穩態對特定波數的擾動變得不穩定時,系統便選擇出一個特徵波長,形成斑點或條紋。
這套機制在斑馬條紋、貝殼花紋與毛囊間距上獲得實證支持。近年最有力的分子證據之一是小鼠指(趾)數目:Sheth 等人(2012)顯示,Hox 基因劑量調節一個近似圖靈系統的波長,改變 Hox 表現會使多趾出現,且趾間距變化與反應—擴散模型預測一致。圖靈機制因此從純數學猜想,成為形態發生的真實設計原則。
基因調控網絡:把濃度變成決定
形態原梯度提供連續輸入,但細胞命運是離散的。介於兩者之間的是基因調控網絡(GRN)——透過轉錄因子間的活化、抑制與互斥構成的邏輯電路。
最典型的模組是互斥抑制開關(toggle switch):兩個轉錄因子彼此抑制,加上自我活化,使系統具有雙穩態。其動力學常以希爾函數(Hill function)描述某基因被活化的程度:
$$ \text{activation} = \frac{[S]^n}{K^n + [S]^n} $$
其中 $[S]$ 為訊號濃度、$K$ 為半飽和常數、$n$ 為希爾係數。當 $n$ 越大,曲線越接近階梯函數,使基因表現呈現近乎開/關的二元切換。高協同性(high cooperativity)因此是把雜訊四溢的形態原梯度,銳化成清晰邊界的關鍵分子手段。果蠅體節形成中的 gap 基因互相抑制,正是這類網絡把平滑梯度切成銳利條紋的實例。
機械力學:形態發生的物理面
化學訊號決定「做什麼」,但組織要彎折、內陷、延伸,仍須真實的力。原腸化時的腹側細胞透過頂端肌動球蛋白(actomyosin)收縮產生頂端收縮(apical constriction),使細胞由柱狀變楔形,整片上皮便向內凹陷。
組織的變形可用連續介質力學近似為黏彈性材料,其行為由表面張力、黏度與主動應力共同決定。一個有用的無因次量是描述黏性力相對於彈性回復的比值;在胚胎尺度(低雷諾數)下,慣性可忽略,主動產生的應力與黏滯阻力達成準平衡。換言之,形態發生是化學圖案與機械力的雙向耦合:基因表現決定哪裡收縮,而收縮產生的張力又回饋影響基因表現(機械轉導,mechanotransduction),形成閉環。這也是為何單看分子或單看力學,都無法完整解釋形態如何湧現。
深入探討(研究所視角)
當代發育生物學的前沿,建立在「定量、單細胞、力學、即時」四股力量的匯流之上,正把上述古典框架推向可預測的工程化階段。
單細胞與時空體學重寫了命運圖譜。 傳統命運圖譜靠少數標記細胞的譜系追蹤,解析度有限。單細胞 RNA 定序(scRNA-seq)配合 RNA velocity(La Manno 等,2018)——利用未剪接與已剪接 mRNA 比例推估轉錄方向,可在高維表現空間重建分化軌跡與分支點。進一步的譜系條碼技術(如 CRISPR/Cas9 誘發的可累積編輯,GESTALT、scGESTALT)把不可逆的基因組「疤痕」當作分子時鐘,讓同一細胞既揭露其表現狀態、又揭露其家族史。結合空間轉錄體學(如 MERFISH、Visium),研究者得以在保留組織座標的前提下測得每個位置的全基因組表現,直接檢驗形態原梯度與下游 GRN 在真實胚胎中的對應,而非僅止於模型推論。
從反應—擴散到 geometry-sensing。 圖靈框架近年被「域幾何回饋」擴充:不只化學決定圖案,組織生長改變的邊界形狀又反過來選擇可容許的波模(Waddington 地景的動態版本)。在合成生物學中,研究者已用合成 Notch 受體(synNotch)與接觸依賴訊號,從零打造能自我組織成多層結構的人工「合成胚胎」,驗證最少多少調控元件足以產生穩定模式。這把 Wolpert 的位置資訊與 Turing 的自組織,從互斥假說整合為連續光譜。
類器官與合成胚胎逼近形態發生的可重製性。 由人類多能幹細胞自組織而成的腦、腸、視杯類器官,以及不需受精的合成胚胎模型(如 ETX、人類 blastoid),讓研究者得以在體外重演原腸化前後的對稱破缺。關鍵發現是這些系統會自發產生 BMP–WNT–NODAL 訊號梯度,重現原本以為非得有母體環境才能建立的軸向資訊——意味著軸向極性可由細胞群體的內在動力學湧現。
機械—化學耦合的結構與系統觀。 在分子尺度,冷凍電鏡解析了 WNT–Frizzled、SHH–PTCH、TGF-β 受體複合體的結構,闡明形態原如何被「呈遞」與運輸(如脂質修飾的 SHH 透過 Dispatched 與脂蛋白顆粒長程傳遞),把訊號傳遞的生化常數錨定到原子結構。在系統尺度,光遺傳學(optogenetics)已能以光在活胚胎中時空精準地開關 ERK 或 Rho 訊號,將過去只能觀察的相關性轉為可操控的因果實驗:人為施加一條梯度,觀察組織是否依模型預測重排。發育生物學由此正從描述性學科,邁向可被定量模型預測、並以光與合成迴路即時編程的物理—資訊科學。