
感覺受器與腦的資訊處理:從轉導動力學到貝氏知覺
放大、編碼與重建——感覺系統如何把物理世界翻譯成神經貨幣
從刺激到知覺:感覺轉導的物理化學界面
當光子打在視網膜、空氣壓力波撞擊耳蝸、或一個葡萄糖分子落在味蕾上,神經系統面對的核心問題只有一個:如何把這些異質的物理化學能量,全部翻譯成同一種「通用貨幣」——膜電位的變化與動作電位的時序碼。這個翻譯過程稱為感覺轉導(sensory transduction),是所有感覺系統共享的第一性原理。本篇不再停留在「感覺受器接收刺激」的層次,而是直接拆解受器分子如何放大、量化並編碼資訊,以及大腦如何重建外在世界。
感覺轉導的本質是一個能量轉換器,把不同形式的物理量(光、機械力、化學濃度、溫度)轉成跨膜離子流。其驚人之處在於增益:一個視桿細胞可偵測單一光子,一個毛細胞能感測小於氫原子直徑的基底膜位移(約 0.3 nm)。要達到這種靈敏度,演化發展出兩條截然不同的策略——離子通道的直接機械門控,以及 G 蛋白偶聯受器(GPCR)驅動的酵素級聯放大。
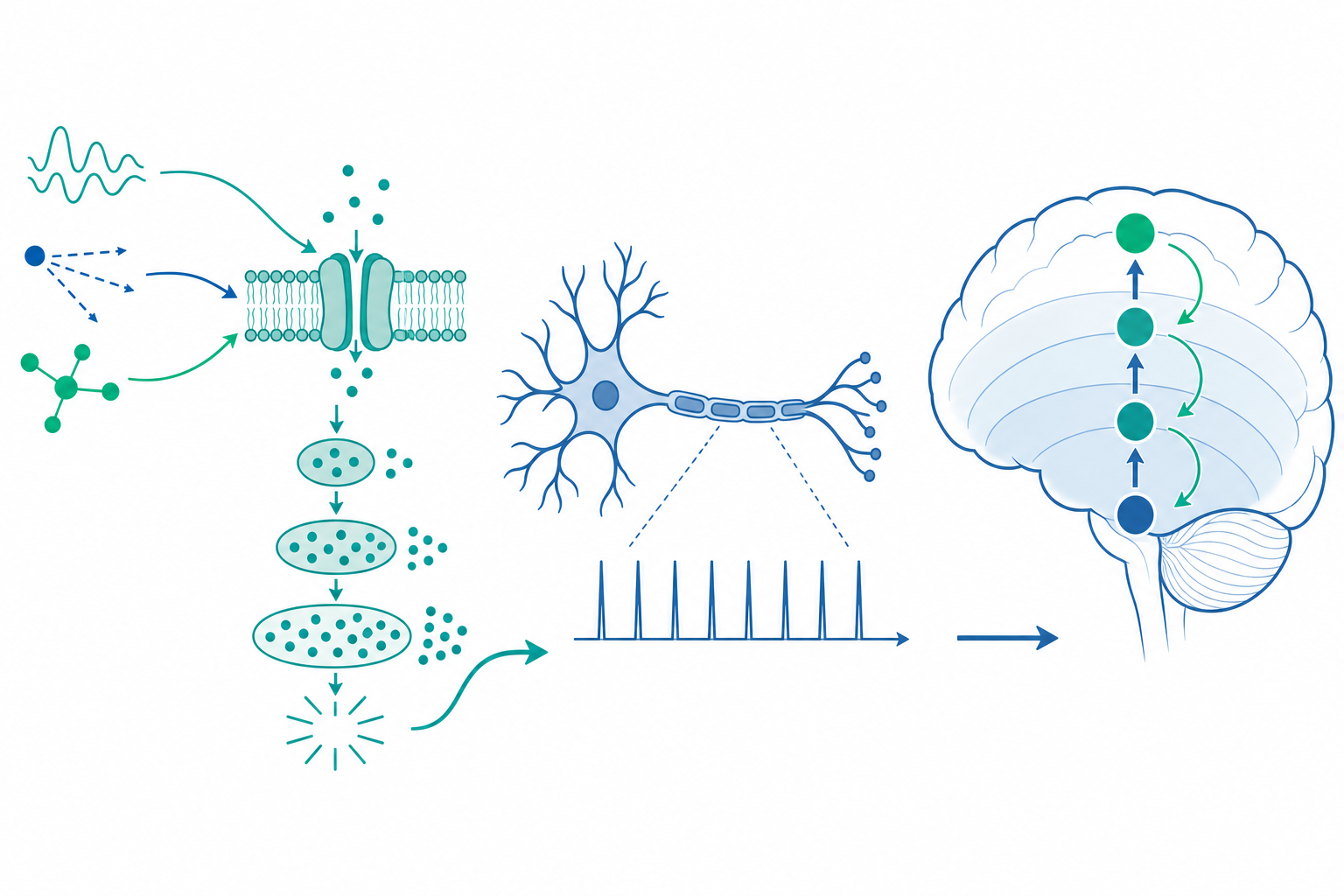
兩種放大策略:直接門控 vs. 級聯放大
機械轉導走的是速度路線。耳蝸毛細胞頂端的纖毛束(stereocilia)由「尖端連結(tip link)」串連,連結蛋白為 cadherin-23 與 protocadherin-15。當基底膜震動使纖毛偏折,尖端連結張力直接拉開機械門控通道(TMC1/TMC2),離子在數十微秒內湧入。這種直接耦合沒有第二信使的延遲,是聽覺能編碼高達數十 kHz 頻率的物理基礎。
光轉導與嗅覺則走放大路線。以視桿為例,一個視紫質(rhodopsin)分子吸收光子後構形改變,活化數百個 transducin(一種 G 蛋白),每個 transducin 活化的 PDE6 又水解上千個 cGMP 分子。cGMP 濃度驟降使 CNG 通道關閉,膜電位超極化。整條級聯的總增益可達 $10^5$ 以上,這正是單光子可被偵測的數學基礎。
級聯反應的動力學可用 Michaelis–Menten 方程式描述酵素步驟的反應速率:
$$v = \frac{V_{max}\,[S]}{K_M + [S]}$$
其中 $[S]$ 為受質濃度,$K_M$ 為達到半飽和速率時的受質濃度。當 $[S] \ll K_M$ 時 $v \approx (V_{max}/K_M)[S]$ 呈線性放大;當 $[S] \gg K_M$ 時 $v \to V_{max}$ 飽和。感覺系統的「適應」很大程度就是動態調整 $K_M$ 與 $V_{max}$,使受器在不同背景強度下都能維持工作於最敏感的線性區。
膜電位的物理基礎:能斯特方程式
無論哪種轉導機制,最終都改變跨膜離子的分布,而離子的平衡電位由 Nernst(能斯特)方程式決定:
$$E_{ion} = \frac{RT}{zF}\ln\frac{[\text{ion}]_{out}}{[\text{ion}]_{in}}$$
其中 $R$ 為氣體常數、$T$ 絕對溫度、$z$ 離子價數、$F$ 法拉第常數。在 37°C 下 $RT/F \approx 26.7$ mV,換成以 10 為底的對數,$K^+$($z=+1$)的平衡電位約為:
$$E_K = 61.5 \times \log_{10}\frac{[K^+]_{out}}{[K^+]_{in}} \text{ mV}$$
定量小範例:耳蝸內淋巴的特殊之處在於 $K^+$ 濃度極高(約 150 mM),而毛細胞內也約 150 mM,但內淋巴維持約 +80 mV 的內淋巴電位。這使得頂端的 $K^+$ 驅動力 = 內淋巴電位(+80 mV)− 毛細胞靜止電位(約 −45 mV)≈ 125 mV 的巨大電化學梯度。正是這個由 stria vascularis 主動維持的「電池」,讓機械門控通道一開啟就有強大的 $K^+$ 內流,賦予聽覺極高的時間解析度。若以 Nernst 計算單純濃度項,當 $[K^+]_{out}/[K^+]_{in}=1$ 時濃度貢獻為 0 mV,故驅動力幾乎全來自那 +80 mV 的電位差——這是一個少見的「以電位而非濃度梯度供能」的例子。
從強度到頻率:感覺編碼的對數律
受器把刺激強度編碼為動作電位的發放頻率。經典的 Weber–Fechner 律指出,主觀感覺強度 $P$ 與刺激強度 $I$ 呈對數關係:
$$P = k\ln\frac{I}{I_0}$$
其中 $I_0$ 為偵測閾值。這解釋了為何聲音強度用分貝(對數單位)、為何我們能同時感知微光與烈日。對數壓縮把橫跨數個數量級的物理刺激,塞進神經元有限的動態範圍(受不應期限制,發放率上限約數百 Hz)。現代研究更傾向 Stevens 冪次律 $P = k I^n$,對不同感覺模態 $n$ 值不同(如電擊 $n \approx 3.5$、亮度 $n \approx 0.33$),但兩者都揭示同一個原則:感覺系統不做線性測量,而是壓縮與正規化。
大腦的資訊處理:群體編碼與預測性重建
訊息進入中樞後,大腦並非被動接收,而是主動推論。在初級感覺皮質,神經元具有感受野(receptive field),且呈拓撲映射(視網膜對應圖、耳蝸頻率圖、體感小人圖)。Hubel 與 Wiesel 的經典工作揭示視覺皮質的方位選擇性柱狀結構,說明大腦透過分層特徵抽取,從點狀光感建構邊緣、輪廓乃至物件。
更深一層,當代神經科學以貝氏推論(Bayesian inference)框架理解知覺:
$$P(\text{world}\mid\text{sense}) \propto P(\text{sense}\mid\text{world})\,P(\text{world})$$
知覺是「先驗信念」與「感覺證據」的最佳整合。錯覺、適應與多感官整合,都是這個推論機制的可預測後果。預測編碼(predictive coding)理論進一步主張:皮質階層自上而下傳遞預測,自下而上傳遞「預測誤差」,只有無法被預測的訊號才會向上傳播——這是一種極度節能的資訊壓縮策略,與感覺受器層次的對數壓縮在精神上一脈相承。
深入探討(研究所視角)
結構生物學揭露轉導的原子細節。 冷凍電子顯微術(cryo-EM)近年徹底改寫了我們對感覺通道的理解。機械門控通道 TMC1 長期是「最後的暗物質」,直到結構解析才確認其與 TMIE、CIB2 形成複合體,並提出脂質作為門控張力傳遞媒介的假說。同樣地,PIEZO1/PIEZO2 通道的三葉螺旋槳狀結構(由 Coste、Patapoutian 團隊鑑定,後者獲 2021 年諾貝爾生理醫學獎)展示了一個橫跨數十奈米的曲面如何把膜張力轉成構形變化——其曲率「碟狀」設計使局部膜形變被高度槓桿放大。溫度受器 TRPV1 的開放與封閉態 cryo-EM 結構,則首次以原子層級呈現辣椒素與熱如何共同調控孔道擴張。
單細胞與空間體學重繪受器多樣性。 單細胞 RNA 定序(scRNA-seq)顛覆了「一種模態一種受器」的舊觀。背根神經節體感神經元被重新分類為十餘個分子亞型,各自表現不同的離子通道與神經肽組合,對應癢、機械痛、溫度等子模態。視網膜亦被解析出超過 40 種神經節細胞型別,每種抽取不同的視覺特徵(運動方向、邊緣、局部對比)並平行送往大腦——這支持「視網膜即是一台前處理運算器」的觀點。空間轉錄體學(spatial transcriptomics)更進一步把這些分子身分定位回組織座標,揭示受器在拓撲圖譜中的精細排列。
系統層次的正規化模型。 在運算神經科學,分裂正規化(divisive normalization)被提議為一條跨物種、跨模態的「正典運算(canonical computation)」。其形式為:
$$R_i = \frac{D_i^{\,n}}{\sigma^{n} + \sum_j D_j^{\,n}}$$
其中 $R_i$ 為第 $i$ 個神經元反應,$D_i$ 為其驅動輸入,分母對鄰近神經元群體加總。此式同時解釋了對比正規化、注意力調節、與多感官整合中的次線性疊加,把前述 Weber–Fechner 對數壓縮統一在一個動態增益控制框架下。
主動感知與閉環。 最前沿的觀點強調感覺絕非單向輸入:眼動、嗅探(sniffing)、觸鬚擺動(whisking)等主動取樣行為,使感覺輸入本身受運動指令調制。皮質中的伴隨放電(corollary discharge)訊號讓大腦區分「自身動作造成的感覺變化」與「外界真實變化」,這也解釋了為何我們無法搔自己癢。把轉導動力學、群體編碼與運動閉環整合進單一生成模型,正是當代「主動推論(active inference)」研究試圖回答的核心問題。