
血紅素與氧解離曲線:協同性的量化機制
從 Hill 方程到 MWC 模型——解讀 S 形曲線背後的分子對話
從「協同合作」談起:氧氣為什麼不平均分配?
如果氧氣與血紅素的結合像一般化學反應那樣「線性比例」,那麼在組織端(低氧分壓)想釋放氧氣時,肺端就會釋放掉太多;想在肺端抓滿氧氣,組織端就抓得太緊。生命選擇了一個更聰明的解:讓血紅素的四個結合位點彼此「商量」。一個位點結合氧氣後,會讓其餘位點更容易結合——這就是協同性(cooperativity),也是氧解離曲線呈 S 形(sigmoidal)而非雙曲線的根本原因。本篇要做的,是把這條曲線從「形狀描述」推進到「可量化的機制模型」。
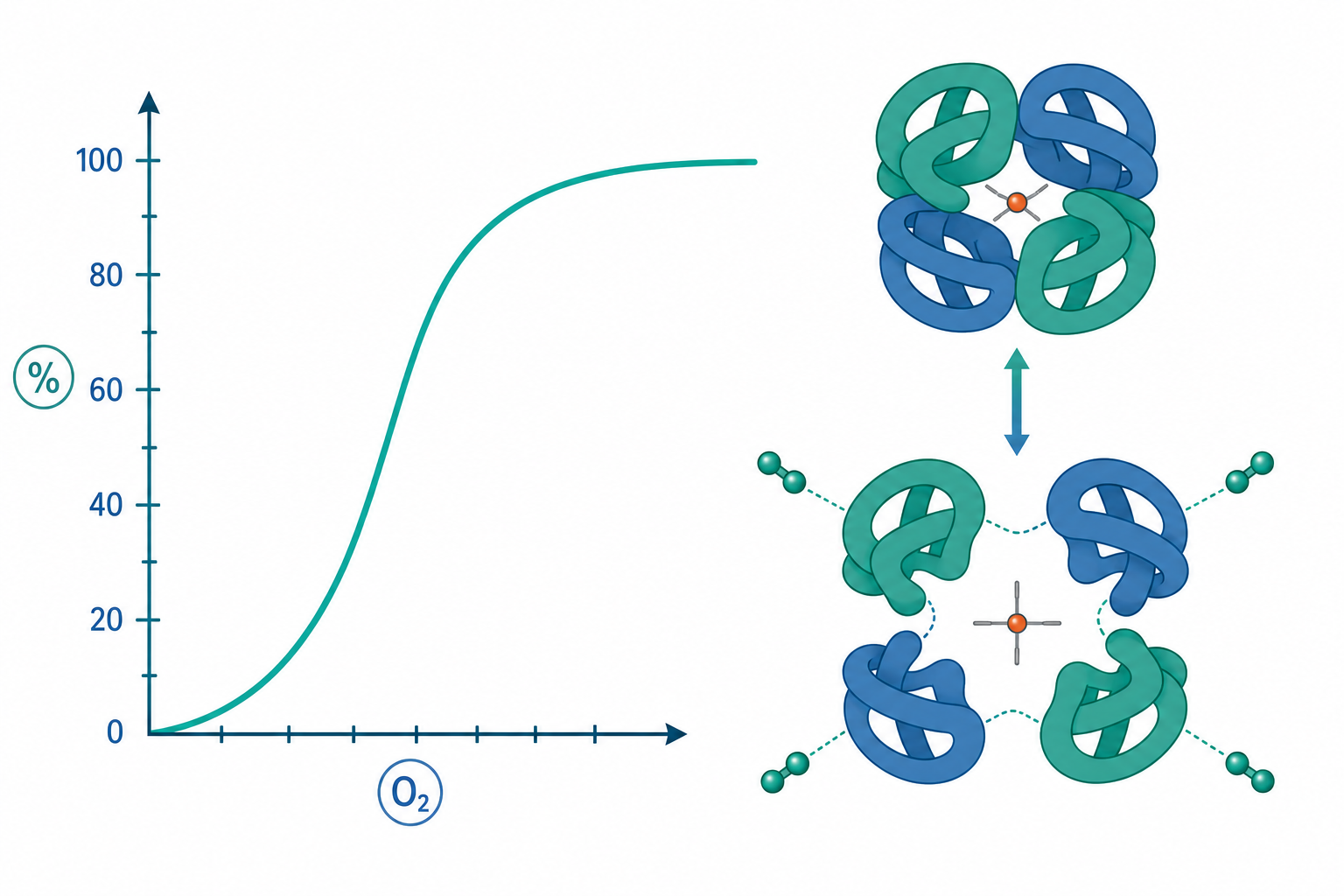
血紅素的四級結構與兩種狀態
成人血紅素(HbA)是 $\alpha_2\beta_2$ 四聚體,每個次單元含一個血基質(heme),其中央 $\mathrm{Fe^{2+}}$ 提供第六配位給氧分子。關鍵在於:血紅素具有兩種可互換的四級結構——緊張態(T state,低氧親和力)與鬆弛態(R state,高氧親和力)。
未結合氧時,血紅素傾向 T 態;當氧分子結合到某個次單元,$\mathrm{Fe^{2+}}$ 從略凸出血基質平面被拉回平面內,牽動近端組胺酸(proximal histidine, F8)與 F 螺旋,進而擾動次單元間的鹽橋,使整個四聚體更傾向 R 態。R 態下其餘位點的氧親和力可提升一到兩個數量級。這就是「結構傳遞訊息」的協同機制。
對單體肌紅蛋白(myoglobin)而言,沒有次單元間的對話,其結合曲線是標準雙曲線(hyperbolic),親和力遠高於血紅素——這正解釋了為何肌肉中的肌紅蛋白能從血液「接手」氧氣。
Hill 方程:把協同性寫成數字
描述氧飽和度 $Y$(已結合位點佔總位點比例)對氧分壓 $P_{O_2}$ 的依賴,最經典的唯象模型是 Hill 方程:
$$Y = \frac{P^{n}}{P_{50}^{\,n} + P^{n}}$$
其中 $P$ 為氧分壓,$P_{50}$ 是達到半飽和($Y=0.5$)所需的氧分壓,$n$(Hill 係數)量化協同程度。將上式整理可得線性化的 Hill plot:
$$\log\!\left(\frac{Y}{1-Y}\right) = n\log P - n\log P_{50}$$
以 $\log[Y/(1-Y)]$ 對 $\log P$ 作圖,斜率即 Hill 係數 $n$。對成人血紅素,實測 $n \approx 2.8$(介於 1 與 4 之間),而非理論上完全協同的 4——這提醒我們:$n$ 並非結合位點數,而是「協同性的有效度量」。$n=1$ 表示無協同(如肌紅蛋白),$n>1$ 表示正協同。
生理上,HbA 的 $P_{50}\approx 26\ \mathrm{mmHg}$。肺泡端 $P_{O_2}\approx 100\ \mathrm{mmHg}$ 時 Hb 幾乎全飽和($Y\approx 0.97$);組織端 $P_{O_2}\approx 40\ \mathrm{mmHg}$ 時 $Y$ 降到約 0.75。正是 S 形曲線在 20–60 mmHg 區間的陡峭斜率,讓血紅素能在組織端「大方卸貨」。
定量小範例:用 Hill 方程算飽和度
已知 HbA 的 $P_{50}=26\ \mathrm{mmHg}$、$n=2.8$,求組織端 $P_{O_2}=40\ \mathrm{mmHg}$ 時的飽和度 $Y$。
先算冪次項。$\left(\dfrac{P}{P_{50}}\right)^{n}=\left(\dfrac{40}{26}\right)^{2.8}=(1.538)^{2.8}$。
取對數:$2.8 \times \log_{10}(1.538) = 2.8 \times 0.187 = 0.524$,故 $(1.538)^{2.8}=10^{0.524}\approx 3.34$。
代入:
$$Y=\frac{(P/P_{50})^{n}}{1+(P/P_{50})^{n}}=\frac{3.34}{1+3.34}\approx 0.77$$
即約 77% 飽和。對照肺端 $P_{O_2}=100\ \mathrm{mmHg}$:$(100/26)^{2.8}\approx 47$,$Y\approx 47/48\approx 0.98$。兩端飽和度差約 21 個百分點——這就是每趟循環卸給組織的氧氣比例。若改用 $n=1$(無協同)重算,組織端 $Y$ 會升到約 0.61、肺端約 0.79,肺端反而抓不滿、組織端卸不乾淨,凸顯協同性的功能價值。
異位調控:曲線會左右移動
氧解離曲線不是固定的。多個異位效應子(allosteric effector)穩定 T 態,使曲線右移($P_{50}$ 升高、親和力下降,利於組織端釋氧):
- $\mathrm{CO_2}$ 與 $\mathrm{H^{+}}$(波耳效應, Bohr effect):組織代謝旺盛處 $\mathrm{CO_2}$ 高、pH 低,質子與胺基末端、組胺酸殘基作用穩定 T 態,促進釋氧;到了肺端反向操作促進結合(Haldane effect 為其互補面)。
- 2,3-雙磷酸甘油酸(2,3-BPG):紅血球糖解副產物,結合於 $\beta$ 鏈間中央正電空腔,僅與 T 態結合。高海拔適應、貧血、慢性缺氧時 2,3-BPG 升高,曲線右移利於組織供氧。
- 溫度:運動產熱使曲線右移。
值得一提的胎兒血紅素(HbF, $\alpha_2\gamma_2$):$\gamma$ 鏈中央腔對 2,3-BPG 親和力低,使 HbF 的 $P_{50}$ 較低(曲線左移、親和力較高),讓胎兒得以從母體血液「奪取」氧氣。這是同一套 Hill 框架下,靠改變 $P_{50}$ 而非 $n$ 達成的生理分流。
兩種協同模型:MWC 與 KNF
Hill 方程雖好用,卻把協同視為「全有全無」的單一步驟,物理上並不真實。兩個更嚴謹的統計力學模型分庭抗禮:
- 協同對稱模型(MWC, Monod–Wyman–Changeux):假設四聚體整體在 T 與 R 兩態間平衡(保持對稱),配體結合改變兩態的平衡常數 $L$。協同性來自「配體偏好 R 態,從而把平衡推向 R」。
- 序列模型(KNF, Koshland–Némethy–Filmer):配體結合誘導該次單元構象改變(誘導契合),再依次影響鄰近次單元,容許中間態。
真實血紅素介於兩者之間,但 MWC 因能優雅地用 $L$、$c$(兩態親和力比)等少數參數重現 S 形曲線與異位效應,至今仍是教科書主力框架。
深入探討(研究所視角)
當代血紅素研究早已超越「兩態切換」的簡化圖像,進入結構動力學、定量配體流動與系統整合的層次。
結構生物學的高解析驗證。 Perutz 以 X 光晶體學解出 T 與 R 兩態結構(1960 年代起),奠定異位機制的原子基礎,這一脈絡為他贏得諾貝爾獎。然而後續以時間解析晶體學(time-resolved crystallography)與低溫電子顯微鏡,配合 Laue 繞射與飛秒 X 光自由電子雷射(XFEL)的研究顯示,配體解離後的構象傳遞並非瞬間完成的二元跳變,而是經過一系列中間態。更精細的工作甚至提出 R、R2 等多個鬆弛態構象,意味著「T↔R 兩態」是一個有用但不完備的近似。核磁共振(NMR)與氫–氘交換質譜進一步揭示,溶液中的血紅素具有顯著的構象異質性與動態漲落,協同性部分根源於構象系綜的重新加權(conformational selection),而非單純的誘導契合——這把 MWC 與 KNF 之爭重新框定為「同一構象景觀的不同投影」。
配體進出的分子路徑。 血基質口袋並非開放於溶劑,氧氣、CO、NO 需穿越蛋白基質。分子動力學模擬與時間解析光譜揭示遠端組胺酸(distal histidine, E7)門控(gating)以及蛋白內部疏水空腔構成的暫態通道,調控配體的進入率與「再結合 vs 逃逸」分配。這對理解 CO 中毒(CO 親和力遠高於 $O_2$)、以及血紅素作為 NO 清除與運輸者的生理角色至關重要——NO–血紅素交互作用牽涉 $\beta$ 鏈 Cys93 的 S-亞硝基化(S-nitrosylation),把氣體訊號傳遞與氧運輸耦合在同一蛋白上。
從單分子到系統與體學。 紅血球層級,單細胞與微流體技術可量測個別細胞的氧釋放動力學異質性,揭示傳統群體平均所掩蓋的分佈。系統生物學則把 Hill/MWC 模型嵌入更大的氧氣輸送方程:結合 $O_2$ 在血漿與組織中的擴散(Krogh 圓柱模型)、微循環的非均勻灌流、以及 $\mathrm{CO_2}$–碳酸氫根緩衝,建立全血氧含量方程 $C_{O_2}=1.34\times[\mathrm{Hb}]\times Y + 0.003\times P_{O_2}$,量化攜帶氧與溶解氧的相對貢獻。臨床上,這些模型支撐了從高海拔生理、鐮刀型紅血球症(單一 $\beta$6 Glu→Val 突變改變聚合行為)到人工血液代用品(cell-free hemoglobin 易引發 NO 清除而升壓)的設計與評估。將分子尺度的協同參數一路接到器官尺度的氧供需平衡,正是當前定量生理學最具整合力的方向。