
不喝水也能喝水:植物如何把水送上百公尺高的樹梢
從蒸散、氣孔到植物激素,看一套不靠幫浦與大腦的生命機器如何運作
植物為什麼不用喝水也能「喝水」?
我們渴了會去倒杯水喝,但一棵高達數十公尺的大樹,既沒有嘴巴,也沒有心臟幫忙打水,它是怎麼把地底下的水送到最高處的葉子的?這個看似簡單的問題,其實藏著植物生理學最迷人的核心:植物是一套不靠肌肉、不靠幫浦,純粹靠物理與化學巧思運作的「生命機器」。
植物的身體大致分成三個工作站:根負責「吸」,莖負責「運」,葉負責「用」。它們之間靠兩條「管路系統」串連——木質部(xylem)負責由下往上輸送水分與礦物質,韌皮部(phloem)負責把葉子製造的養分送往全身。你可以把木質部想成「上水道」,韌皮部想成「外送服務」。理解這兩條管路怎麼運作,就理解了植物如何生活。
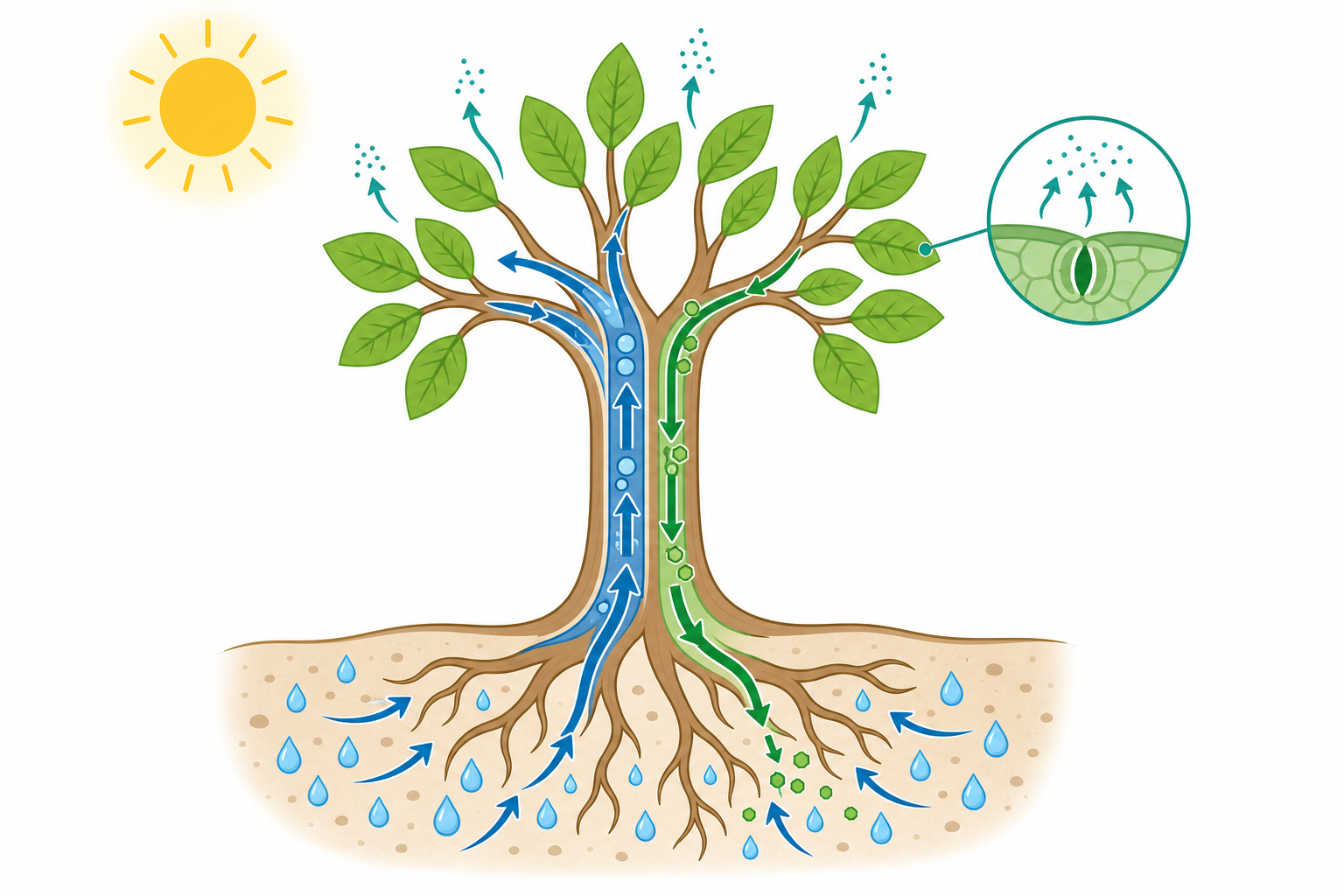
蒸散作用:太陽才是真正的抽水馬達
水往高處走,似乎違反直覺,畢竟地心引力是往下拉的。植物的祕訣是:它根本不「推」水上去,而是從頂端「拉」上去。
關鍵在於蒸散作用(transpiration)。葉片表面有許多小孔叫氣孔(stomata),水分會從這裡蒸發到空氣中。當頂端的水分子離開時,因為水分子之間有強烈的內聚力(cohesion)——彼此像手牽手一樣——一個離開就會把下一個拉上來,形成一條從根到葉、連續不斷的「水柱」。這就是著名的內聚力-張力學說(cohesion-tension theory)。
打個比方:這就像你用一根長吸管喝珍珠奶茶,你在上面吸,整條飲料就被往上拉。差別只在於,植物的「吸力」來源是太陽——是陽光蒸發了葉子的水,免費替植物抽了一整天的水。
這個力量大得驚人。水柱承受的張力可以讓水爬升超過 100 公尺(紅杉就是活生生的例子)。同時,水分子與木質部細小管壁之間還有附著力(adhesion),幫忙抵抗下滑,這兩股力量合稱毛細現象。
氣孔的兩難:要喝水,還是要呼吸?
氣孔不只是讓水跑掉的破洞,它更是植物的呼吸窗口。光合作用需要二氧化碳($\mathrm{CO_2}$),而 $\mathrm{CO_2}$ 正是從氣孔進來的。
這就形成一個經典的兩難:氣孔開著,可以吸進 $\mathrm{CO_2}$ 行光合作用,但水分也會大量蒸散流失;氣孔關著,保住了水,卻也餓著肚子。 植物每天都在做這道權衡題。
控制氣孔的是兩個香腸狀的保衛細胞(guard cells)。當它們吸水膨脹,氣孔張開;失水萎縮,氣孔閉合。在炎熱乾旱的中午,植物常會主動關閉部分氣孔來保命——這也是為什麼盛夏正午的植物看起來「無精打采」。
光合作用與呼吸作用的總反應式,正好是一對相反的化學方程式:
$$6\,\mathrm{CO_2} + 6\,\mathrm{H_2O} \xrightarrow{\text{光能}} \mathrm{C_6H_{12}O_6} + 6\,\mathrm{O_2}$$
$$\mathrm{C_6H_{12}O_6} + 6\,\mathrm{O_2} \longrightarrow 6\,\mathrm{CO_2} + 6\,\mathrm{H_2O} + \text{能量}$$
光合作用把太陽能「存」進葡萄糖,呼吸作用再把它「提領」出來供應生命活動——植物白天主要做前者,但其實兩者整天都在進行。
韌皮部:糖的雙向外送系統
葉子用光合作用做出糖之後,要送到不會行光合作用的地方,例如根、果實、生長中的嫩芽。這條外送路線就是韌皮部,運作原理稱為壓力流學說(pressure-flow hypothesis)。
我們用一組「來源—去處」的對照來說明:
- 來源(source):通常是葉子。糖被主動運入韌皮部,使該處糖濃度升高,水因滲透作用跟著流入,產生高壓。
- 去處(sink):例如正在長大的果實。糖被卸下利用,濃度降低,水流出,壓力變低。
於是韌皮部裡形成「高壓推向低壓」的流動,把糖水從來源一路擠到去處。妙的是,韌皮部的方向不固定:春天樹根把儲存的養分往上送給新芽,此時根是來源、芽是去處;夏天葉子把糖往下送給根儲存,方向就反過來了。這正是韌皮部「雙向外送」的彈性所在。
植物激素:沒有大腦的精準指揮
植物沒有神經系統,卻能精準地朝光彎曲、讓果實成熟、在秋天落葉。指揮這一切的是化學信使——植物激素(plant hormones)。
最著名的例子是生長素(auxin)造成的「向光性」。當光從側面照來,生長素會累積到背光的那一側,促使該側細胞伸長得比向光側快,於是莖就「彎向」光源——這不是植物在「看」光,而純粹是激素分布不均的結果。
其他重要激素還有:促進果實成熟與落葉的乙烯(ethylene,一種氣體),這也是為什麼把熟蘋果和生香蕉放一起,香蕉會熟得更快;幫助種子打破休眠、莖節伸長的激勃素(gibberellin);以及在乾旱時下令關閉氣孔、堪稱「壓力荷爾蒙」的離層酸(ABA)。這些激素彼此拮抗或協同,組成一套沒有大腦卻無比靈巧的調控網路。
深入探討(研究所視角)
入門段把蒸散流描述為連續水柱,但這條水柱其實處於負壓(張力)狀態,這在熱力學上是個亞穩態,極易發生空穴化(cavitation)——水柱因張力過大而斷裂、形成氣泡,導致木質部導管栓塞(embolism)失去輸水能力。植物在乾旱或結凍時尤其脆弱。現代生理學以易損性曲線(vulnerability curve)量化此風險,並以水力構造的「安全邊際」概念解釋為何不同物種在乾旱下存活率迥異。新近研究更發現,部分植物能透過韌皮部分泌與根壓在夜間「修復」栓塞,這牽涉到木質部薄壁細胞主動分泌的滲透調節,是當前水力生理學的熱點。
在分子層次,氣孔調控已被解析得相當細緻。離層酸(ABA)與其受體 PYR/PYL/RCAR 結合後,抑制蛋白磷酸酶 PP2C,解放激酶 SnRK2,再活化保衛細胞質膜上的離子通道(如陰離子通道 SLAC1),驅動鉀離子與陰離子外流、保衛細胞失水閉孔。這條 PYR–PP2C–SnRK2 訊息軸是利用 CRISPR/Cas9 進行抗旱作物改良的核心標靶;研究者藉由編輯受體或通道基因,調校作物在缺水下的氣孔反應與水分利用效率(WUE)。
生長素的極性運輸同樣有精緻的分子基礎:PIN 蛋白家族不對稱地分布在細胞膜的特定一側,像單行道路標一樣決定生長素的流向,這套「化學滲透極性運輸模型」由 Rubery、Sheldrake 與 Goldsmith 等人奠基,是植物形態建成(morphogenesis)的關鍵。
方法學上,單細胞 RNA 定序(scRNA-seq)正在重寫我們對根與葉發育的理解——過去只能談「組織」,如今能逐一細胞描繪轉錄狀態圖譜,揭示保衛細胞、伴細胞、根毛細胞各自的命運決定軌跡。結合系統生物學的網路模型,研究者得以把激素訊號、轉錄調控與水力物理整合進同一套動力學框架,預測植物在氣候變遷下的生理極限。這些工具也連結到更大的議題:在 $\mathrm{CO_2}$ 濃度升高、極端乾旱頻發的世界裡,理解氣孔的開閉邏輯,正是預測糧食安全與碳匯能力的關鍵一環。