
CRISPR–Cas9 基因編輯:從分子辨識邏輯到動力學與系統視角
以 PAM/R-loop 辨識、切割動力學與修復途徑競爭,剖析可程式化基因編輯的機制與前沿應用
從細菌免疫到精準分子手術刀
當你聽到「CRISPR–Cas9 是基因剪刀」時,這個比喻其實掩蓋了一個更精緻的真相:它本質上是一套可程式化的序列辨識酵素系統,由 RNA 引導蛋白質在數十億鹼基的基因組中找到唯一的標靶。理解它的威力,必須從動力學、熱力學與酵素學的層次切入,而不只是停留在「剪斷 DNA」的圖像。
CRISPR(Clustered Regularly Interspaced Short Palindromic Repeats)原本是細菌與古菌對抗噬菌體的適應性免疫系統。當噬菌體入侵後,細菌會將一小段病毒 DNA(spacer,約 20–40 bp)插入自身基因組的 CRISPR 陣列;再次遭遇時,這段記憶被轉錄為 crRNA,與 Cas 蛋白組成複合體,精準切割入侵者的核酸。Doudna 與 Charpentier 在 2012 年的關鍵工作,將天然系統中的 crRNA 與 tracrRNA 融合成單一的 single-guide RNA(sgRNA),把這套防禦機制改造成可在任意物種中編程的工具。
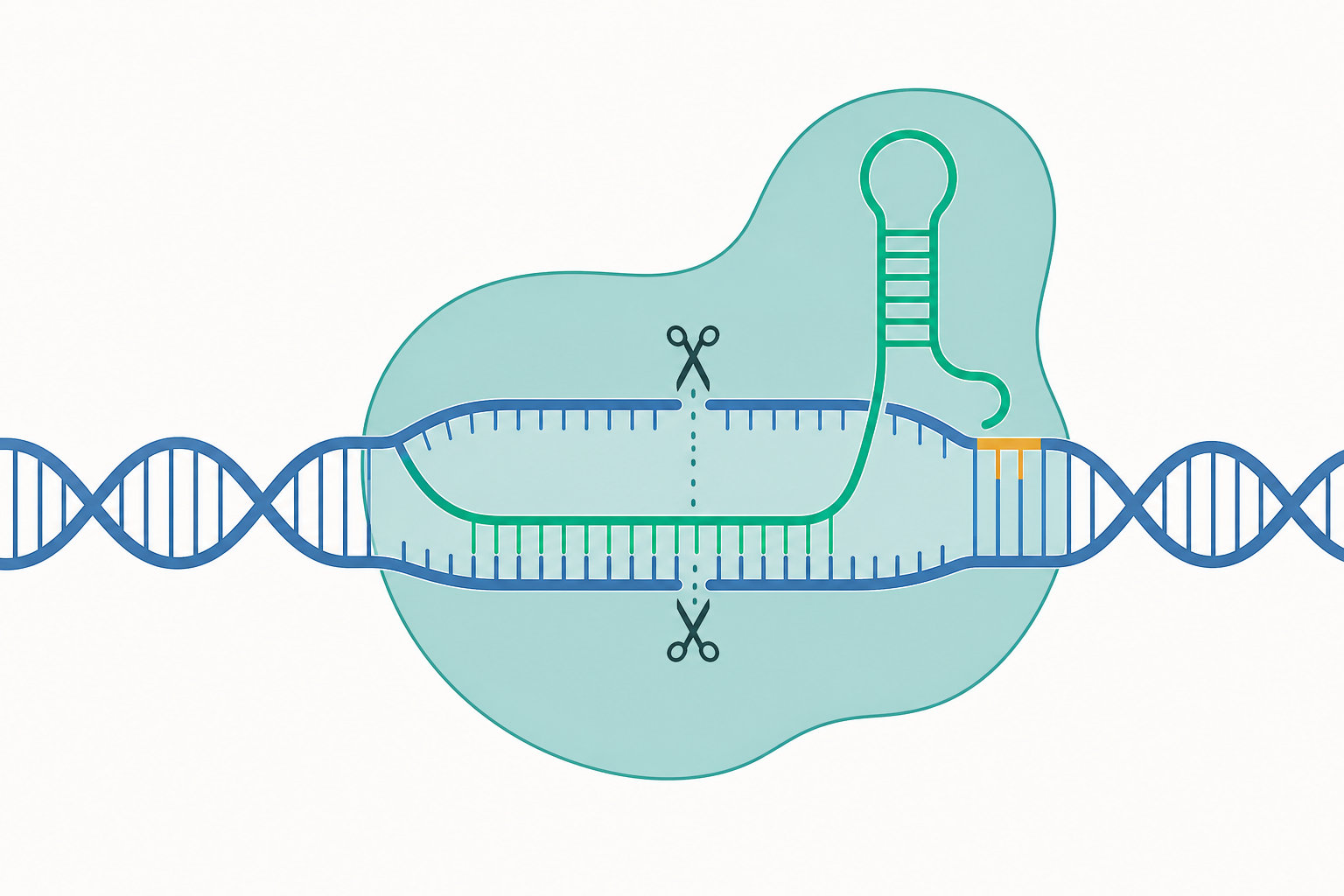
標靶辨識的分子邏輯:PAM 與 R-loop
Cas9 的辨識並非單純的鹼基互補。整個過程是一連串具有嚴格次序的構象檢查點(conformational checkpoints)。
第一道關卡是 PAM(Protospacer Adjacent Motif)。對 Streptococcus pyogenes 的 SpCas9 而言,PAM 序列為 5′-NGG-3′,位於標靶序列(protospacer)的 3′ 端。Cas9 並非隨機掃描,而是先以一維/三維混合的方式在基因組上快速碰撞,只有當 PAM-interacting domain 辨識到 NGG 時,才會啟動 DNA 雙股的局部解旋。這個設計也解釋了細菌為何不會切到自己的 CRISPR 陣列——因為陣列旁缺乏 PAM。
PAM 確認後,sgRNA 的 spacer 開始與標的股進行鹼基配對,由 PAM 近端往遠端逐步形成 R-loop(RNA-DNA 雜交雙股 + 被置換的非標的單股)。這個過程在熱力學上可粗略視為:
$$\Delta G_{\text{total}} = \Delta G_{\text{RNA:DNA}} - \Delta G_{\text{DNA:DNA}} + \Delta G_{\text{Cas9}}$$
亦即 RNA-DNA 雜交所釋放的自由能,必須足以補償拆開原本 DNA 雙股所需的能量,再加上蛋白質構象變化的貢獻。seed region(PAM 近端約 8–12 nt)的配對最為關鍵:此處一個錯配往往使 R-loop 無法穩定延伸,這是 Cas9 特異性的物理基礎。相對地,PAM 遠端的少數錯配可能被容忍,正是 off-target(脫靶)切割的來源。
只有當 R-loop 完整延伸至 20 nt,HNH 與 RuvC 兩個核酸酶結構域才會發生構象重排而被活化:HNH 切割標的股,RuvC 切割非標的股,在 PAM 上游約 3 bp 處產生鈍端雙股斷裂(DSB)。
切割動力學的量化視角
Cas9 在動力學上是一個典型的單轉換(single-turnover)酵素:切割後它仍緊抓著產物 DNA 數小時,幾乎不釋放。因此用古典的 Michaelis–Menten 方程描述穩態速率:
$$v = \frac{V_{\max}[S]}{K_m + [S]}$$
時,必須謹慎——Cas9 的限速步驟並非化學切割,而是切割「之前」的 R-loop 形成與構象校對,以及切割「之後」的產物釋放。研究中常以 $k_{\text{cleave}}$(切割速率常數)與表觀 $K_d$ 來刻畫不同 sgRNA 的效率。
定量小範例: 假設某 sgRNA 的單分子切割服從一級動力學 $[\text{uncut}]_t = [\text{uncut}]_0\, e^{-k_{\text{obs}} t}$。若實驗測得切割半衰期 $t_{1/2}=30$ 秒,求 $k_{\text{obs}}$ 並估算 90 秒後仍未被切割的比例。
由 $t_{1/2}=\dfrac{\ln 2}{k_{\text{obs}}}$:
$$k_{\text{obs}} = \frac{\ln 2}{30\ \text{s}} \approx \frac{0.693}{30} \approx 0.0231\ \text{s}^{-1}$$
90 秒 = 3 個半衰期,故未切割比例為:
$$\frac{[\text{uncut}]_{90}}{[\text{uncut}]_0} = e^{-0.0231\times 90} \approx e^{-2.08} \approx 0.125 = 12.5\%$$
亦即約 87.5% 的標靶已被切割。這類動力學分析讓我們能客觀比較不同 sgRNA 設計的優劣,而非僅憑終點凝膠的「有切到沒切到」。
斷裂之後:細胞如何決定編輯結果
Cas9 只負責製造 DSB,真正改寫基因組的是細胞自身的 DNA 修復途徑,這是基因編輯結果最大的變數來源。
- 非同源端接合(NHEJ):快速但易出錯,在接點引入隨機的插入或缺失(indel)。若 indel 長度非 3 的倍數,會造成框移突變(frameshift)而使基因失活——這是基因「敲除」(knockout)的主要機制。
- 同源導向修復(HDR):以提供的 DNA 模板為藍本精確修復,可用於「敲入」(knock-in)特定序列。但 HDR 僅在細胞週期的 S/G2 期活躍,效率通常遠低於 NHEJ。
修復途徑的競爭可粗略以機率分配理解:若 $P_{\text{NHEJ}}$ 與 $P_{\text{HDR}}$ 為兩途徑被選用的機率($P_{\text{NHEJ}}+P_{\text{HDR}}\approx 1$,忽略 MMEJ 等次要途徑),則提高精準編輯效率的策略,本質上都是在移動這個機率天平——例如同步抑制 NHEJ 關鍵蛋白、將細胞同步在 S 期、或縮短模板與斷點的距離。
應用版圖與第二代工具
理解了機制,應用便水到渠成。在基礎研究中,CRISPR 讓全基因組規模的功能篩選(pooled screen)成為常規。在治療上,2023 年核准的鐮刀型貧血/β 地中海貧血療法,正是以 ex vivo 編輯造血幹細胞、重新活化胎兒血紅素來達成。
機制上的限制也催生了第二代工具,它們的共同策略是「不依賴 DSB」:
- 鹼基編輯(Base editing):將催化失活的 Cas9(nickase 或 dCas9)與去胺酶融合,直接把 C·G 轉成 T·A 或 A·T 轉成 G·C,無需斷裂雙股,大幅降低 indel 與大片段缺失風險。
- 先導編輯(Prime editing):將 Cas9 nickase 與反轉錄酶融合,由帶有編輯資訊的 pegRNA 引導,理論上可達成所有 12 種點突變與小片段插入缺失,且不依賴 HDR 與外源模板。
深入探討(研究所視角)
主體談的是「平均行為」,但前沿研究的精彩之處,在於把 Cas9 拆解到單分子、結構與系統的時間與空間尺度。
結構生物學與構象校對(conformational proofreading)。 冷凍電顯(cryo-EM)與單分子 FRET 揭示,Cas9 的特異性並非單純由鹼基配對的熱力學決定,而是一種動力學校對。當 R-loop 完整且正確時,HNH 結構域會經歷一段大幅度(約 30 Å)的構象擺動,從一個「校驗位置」移動到緊貼切割位點的「活化位置」;錯配會抬高這個構象轉變的能障,使 HNH 卡在非活化態,DNA 雖被結合卻不被切割。這意味著 Cas9 在化學切割「之後且之前」都設有檢查點——這正是工程化高保真變體(如 HiFi-Cas9、eSpCas9、HypaCas9)的設計原理:在 REC3 等結構域引入突變,刻意升高活化所需的構象門檻,犧牲少許切割速率換取大幅降低脫靶。
單細胞與多體學視角。 大規模功能基因組學已從「敲除後看表型」進化為 Perturb-seq:把 CRISPR 擾動與單細胞 RNA-seq 偶聯,使每個細胞同時帶有「被擾動的基因」標籤與「全轉錄組讀出」。這讓研究者能在單次實驗中建構數千個基因擾動對應的轉錄組反應圖譜,進而重建基因調控網路(GRN)。配合 CRISPRi(dCas9-KRAB 抑制)與 CRISPRa(dCas9-activator 活化),同一個 dCas9 平台可在 Cognomics 之外,於分子層次實現「可程式化的基因表達調控」,而非僅是序列破壞。
系統生物學與脫靶的量化模型。 脫靶預測已從單純的錯配計數,發展為機器學習與生物物理混合模型。其核心仍可回到自由能景觀:每個潛在脫靶位點的被切割機率,取決於該位點 R-loop 能否越過構象活化能障,可用類似波茲曼分布 $P_i \propto e^{-\Delta G_i / k_BT}$ 的權重來排序候選位點。GUIDE-seq、CIRCLE-seq、DISCOVER-seq 等全基因組脫靶定位技術,則提供了校正這些模型的實驗真值(ground truth)。
演化與群體層次的張力。 當 CRISPR 被設計成基因驅動(gene drive)——讓某等位基因以遠高於孟德爾 50% 的比例傳遞時,它直接挑戰了 Hardy–Weinberg 平衡的核心假設(無天擇、無遷移、隨機交配下等位基因頻率不變,$p^2+2pq+q^2=1$)。基因驅動人為製造了極強的傳遞偏倚(transmission distortion),理論上可在數十個世代內把一個等位基因推向固定。這既是控制瘧蚊等病媒的潛在利器,也帶來不可逆改變野生族群的生態與倫理風險——提醒我們:分子尺度的精準,一旦放大到族群與演化尺度,後果的可預測性反而急遽下降。