
從一隻到一群:生物數量如何爆發、煞車與拉鋸
指數與邏輯斯成長、種間競爭、捕食與群落消長的完整圖像
從一隻到一群:生物數量是怎麼變化的
想像你在春天的池塘裡放進幾隻浮萍。一週後,水面開始出現綠點;兩週後,半個池塘被覆蓋;到了某一天,整個池塘綠油油一片,再也擠不下新的浮萍。這個過程其實濃縮了生態學最核心的問題之一:一群生物的數量,到底是怎麼增加、又是怎麼停下來的?
「族群」(population)指的是同一物種、生活在同一地區、能彼此交配繁殖的一群個體。研究族群數量如何隨時間變化,正是「族群生態學」的主軸。而當不同族群擠在同一片棲地、彼此競爭、捕食、共生時,就形成了「群落」(community)。本文就從一隻,講到一群,再講到許多群之間的拉鋸。
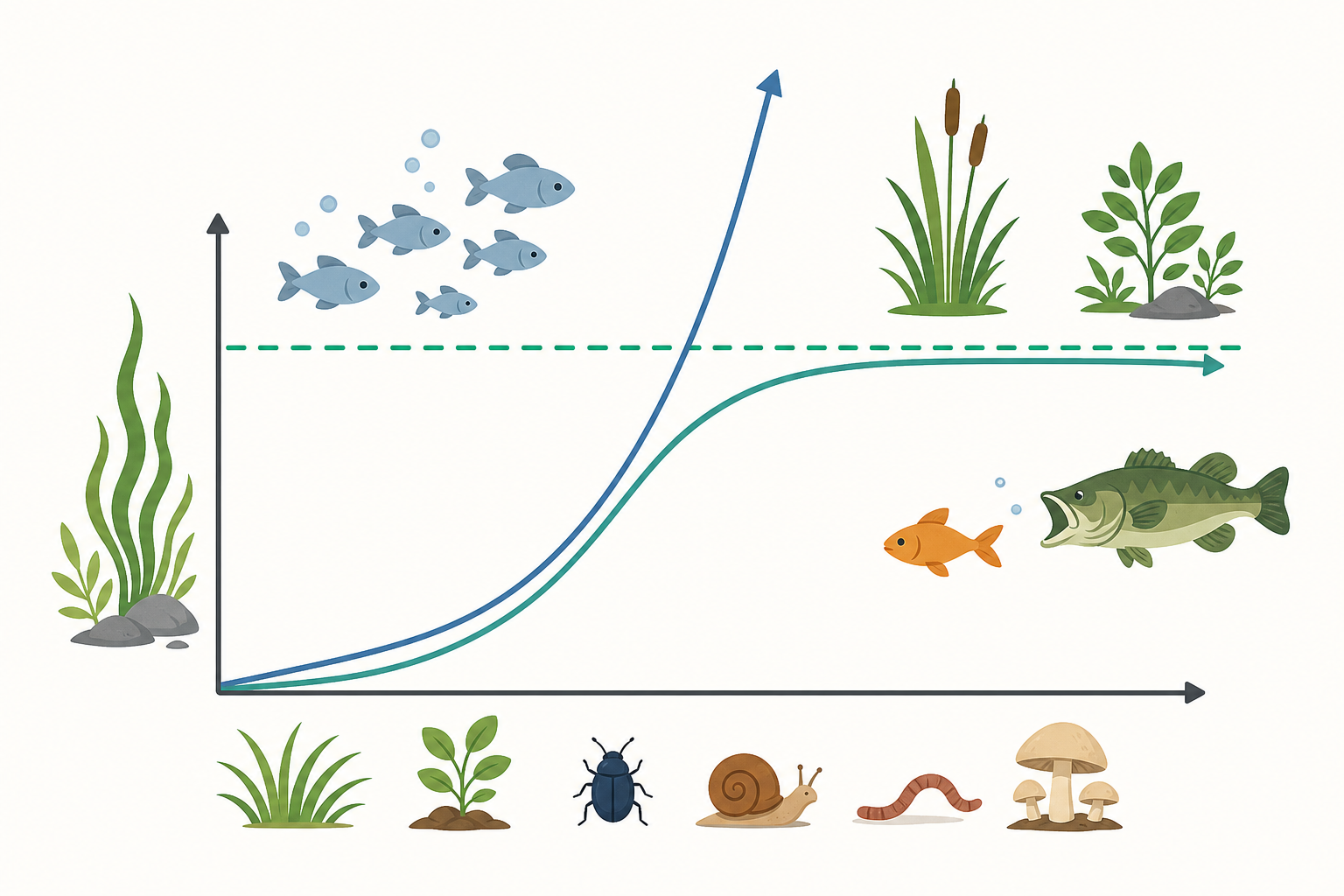
指數成長:理想世界裡的爆炸
如果食物無限、空間無限、沒有天敵也沒有疾病,一個族群會怎麼長?答案是「指數成長」(exponential growth)。每隻個體都盡情繁殖,下一代再繁殖,數量像滾雪球一樣翻倍再翻倍。
我們可以用一條簡單的式子描述族群數量 $N$ 隨時間 $t$ 的變化速率:
$$\frac{dN}{dt} = rN$$
這裡的 $r$ 是「內在增長率」(每個個體平均的淨增加速度),$N$ 是目前的數量。式子讀起來很直白:現有個體越多,增加得越快。畫成圖,是一條越翹越陡的 J 形曲線。
舉個例子:假設一種細菌每 20 分鐘分裂一次。從 1 隻開始,1 小時後有 $2^3 = 8$ 隻,2 小時後是 $2^6 = 64$ 隻,10 小時後就超過十億隻。這就是為什麼食物放久了會「整鍋壞掉」——細菌正在它的理想世界裡爆炸。
但現實中,沒有任何族群能永遠這樣長。池塘會被浮萍塞滿,食物會吃光,廢物會累積。於是我們需要第二條曲線。
邏輯斯成長:撞上環境的天花板
真實環境有極限。生態學家用「環境負載量」(carrying capacity,記為 $K$)來表示某片棲地最多能養活多少個體。當族群數量接近 $K$,資源變少、競爭變激烈,增長就會慢下來,最後停在 $K$ 附近上下擺盪。
把這個「煞車」加進式子,就得到「邏輯斯成長」(logistic growth):
$$\frac{dN}{dt} = rN\left(1 - \frac{N}{K}\right)$$
關鍵在那個括號 $\left(1 - \dfrac{N}{K}\right)$,它扮演剎車的角色:
- 當 $N$ 很小(遠小於 $K$):括號接近 1,式子幾乎等於 $rN$,族群快速成長,像指數那段。
- 當 $N$ 接近 $K$:括號接近 0,增長速率趨近於零,族群停止擴張。
- 當 $N$ 等於 $K$:增長為 0,達到平衡。
畫成圖是一條優雅的 S 形(sigmoid)曲線:先慢、再快、最後又慢下來貼著天花板。許多實驗室培養的酵母菌、草履蟲,都呈現漂亮的 S 形成長。回到開頭的浮萍:它一開始指數爆發,但池塘大小就是它的 $K$,撞到天花板後就鋪滿水面、不再增加了。
種間競爭:一山難容二虎
當兩個物種需要同樣的資源(食物、陽光、築巢點),它們之間就會發生「種間競爭」。俄國生態學家高斯(Gause)用兩種草履蟲做過經典實驗:單獨培養時,兩種都長得很好;但放在同一個試管裡爭奪同樣的食物時,其中一種會逐漸壓過另一種,最後把對方排擠到滅絕。
這帶出了著名的競爭排除原則(competitive exclusion principle):兩個生態棲位(niche)完全相同的物種,無法在同一地方長期共存。要嘛一方勝出,要嘛雙方「分工」——例如一種在白天活動、另一種改成夜間覓食,或一種吃大種子、另一種吃小種子。這種把資源切開分用的現象,叫做「棲位分化」,它讓更多物種得以共存,也是群落多樣性的來源之一。
捕食與消長:你追我跑的舞蹈
群落裡另一組重要關係是「捕食」。捕食者與獵物的數量常常呈現規律的「你追我跑」:獵物多 → 捕食者吃得飽、繁殖變多 → 捕食者多到把獵物吃少 → 捕食者沒食物、數量下降 → 獵物又開始回升。如此循環,兩條曲線像跳探戈一樣交錯起伏。加拿大山貓與雪兔長達近百年的毛皮交易紀錄,就呈現出這種周期性的波動。
而當尺度拉得更大、時間更長,整個群落還會發生「消長」(succession)。一片火災後的裸地,會先長出耐旱的草與雜草(先驅物種),它們改善土壤後,灌木進駐,接著小樹、大樹陸續取代,最後形成穩定的森林(極相群落)。生物數量的變化,從個體、族群,一路串連到整個生態系的緩慢演替。
把這些串起來
從指數成長的 J 形爆炸,到邏輯斯成長的 S 形煞車;從兩物種的競爭排除,到捕食者與獵物的周期舞蹈,再到群落的長期消長——這些都是在回答同一個問題:生物的數量為什麼會這樣變? 理解這些規律,我們才能解釋為什麼入侵種會失控、為什麼保育需要維持最小族群、為什麼一片棲地能養活多少生物有其上限。
深入探討(研究所視角)
入門段落把族群當成單一數字 $N$,但進階研究會把這個數字「拆開」。矩陣族群模型(Leslie matrix)把族群依年齡或發育階段分層,用一個矩陣 $\mathbf{L}$ 描述各階段的存活率與生育率,族群向量隨世代更新為 $\mathbf{n}_{t+1} = \mathbf{L}\,\mathbf{n}_t$。矩陣的「優勢特徵值」$\lambda$ 即長期幾何增長率($\lambda > 1$ 成長、$<1$ 衰退),對應的特徵向量則是穩定的年齡結構。這讓保育生物學能精準分析「保護哪個年齡層最有效」——例如海龜研究指出,提升成龜存活率對族群延續的貢獻,遠大於保護大量的龜卵。
種間競爭的數學骨架是洛特卡—沃爾泰拉方程(Lotka–Volterra equations)。兩物種的競爭可寫成一組耦合微分方程,引入競爭係數 $\alpha_{12}$、$\alpha_{21}$ 表示彼此的抑制強度。透過分析其「零增長等斜線」(zero-growth isoclines)的交點,可推導出四種結局:物種 1 勝、物種 2 勝、不穩定共存、或穩定共存。同樣框架略加修改即可描述捕食者—獵物動態,並解釋前述的周期性振盪。現代族群生態學更進一步引入隨機性:當族群極小時,純粹的運氣(人口統計隨機性,demographic stochasticity)就可能導致滅絕,這是「最小存活族群」(MVP)概念的理論基礎。
分子工具正在改寫族群生態學的尺度。傳統靠標記—再捕捉估計數量,現在則用環境 DNA(eDNA):採一瓶水或一把土,定序其中殘留的生物 DNA,就能偵測哪些物種曾經過、甚至估算相對豐度,對稀有或隱蔽物種尤其有用。族群基因體學(population genomics)以全基因組 SNP 資料估計有效族群大小 $N_e$、近交程度與基因流,揭露「數量看似很多、但遺傳多樣性已瀕危」的隱性風險。此外,metabarcoding 與單一樣本高通量定序讓整個群落的物種組成得以快速盤點,把「群落」從觀察名單變成可量化的分子指紋。這些技術與經典的 $\frac{dN}{dt}$ 模型結合,正讓族群與群落生態學從描述走向預測——預測入侵、預測崩潰、也預測在氣候變遷下哪些族群能撐過下一個世紀。