
植物激素與環境訊號傳遞:從受體動力學到運算網路
以結合常數、降解動力學與離子電化學,量化植物如何讀懂世界
從「分子開關」看植物如何讀懂世界
植物不能逃跑,因此牠們把所有「決策」都壓縮成化學訊號的濃度變化。一個簡單的直覺是:植物激素(phytohormone)就是一群在極低濃度(常為 $10^{-9}$ 至 $10^{-6}$ M)下就能改變細胞命運的小分子。但若停在「生長素促進生長、離層酸抑制生長」這種層次,就會錯過真正精彩的部分——訊號傳遞的本質是受體—配體結合動力學、蛋白質降解速率、與基因表現之間的量化耦合。本文聚焦激素感知與環境訊號整合的分子機制,並用幾個定量模型把直覺收緊成可計算的描述。
激素感知:從受體結合到「降解即訊號」
近二十年植物訊號領域最深刻的轉變,是發現多種激素的核心受體機制其實是操控蛋白質降解,而非典型的第二信使級聯。以生長素(auxin, IAA)為例:受體 TIR1 是一個 F-box 蛋白,IAA 像「分子膠水」一樣坐進 TIR1 與 Aux/IAA 抑制蛋白之間的口袋,促成 SCF$^{\text{TIR1}}$ 泛素接合酶將 Aux/IAA 標記、送進 26S 蛋白酶體降解,從而釋放 ARF 轉錄因子。茉莉酸(JA-Ile)透過 COI1–JAZ、激勃素(GA)透過 GID1–DELLA,都遵循同一個「受體促進抑制子降解」的範式。
把這件事量化,受體—配體的結合可用平衡常數描述:
$$\theta = \frac{[L]}{[L] + K_d}$$
其中 $\theta$ 為受體佔據率,$[L]$ 為自由激素濃度,$K_d$ 為解離常數。TIR1–IAA 的 $K_d$ 量級約在數十至數百 nM,意味著當細胞內 IAA 濃度橫跨此範圍時,受體佔據率對濃度的變化最為敏感——這正是植物把「濃度梯度」翻譯成「反應差異」的物理基礎。
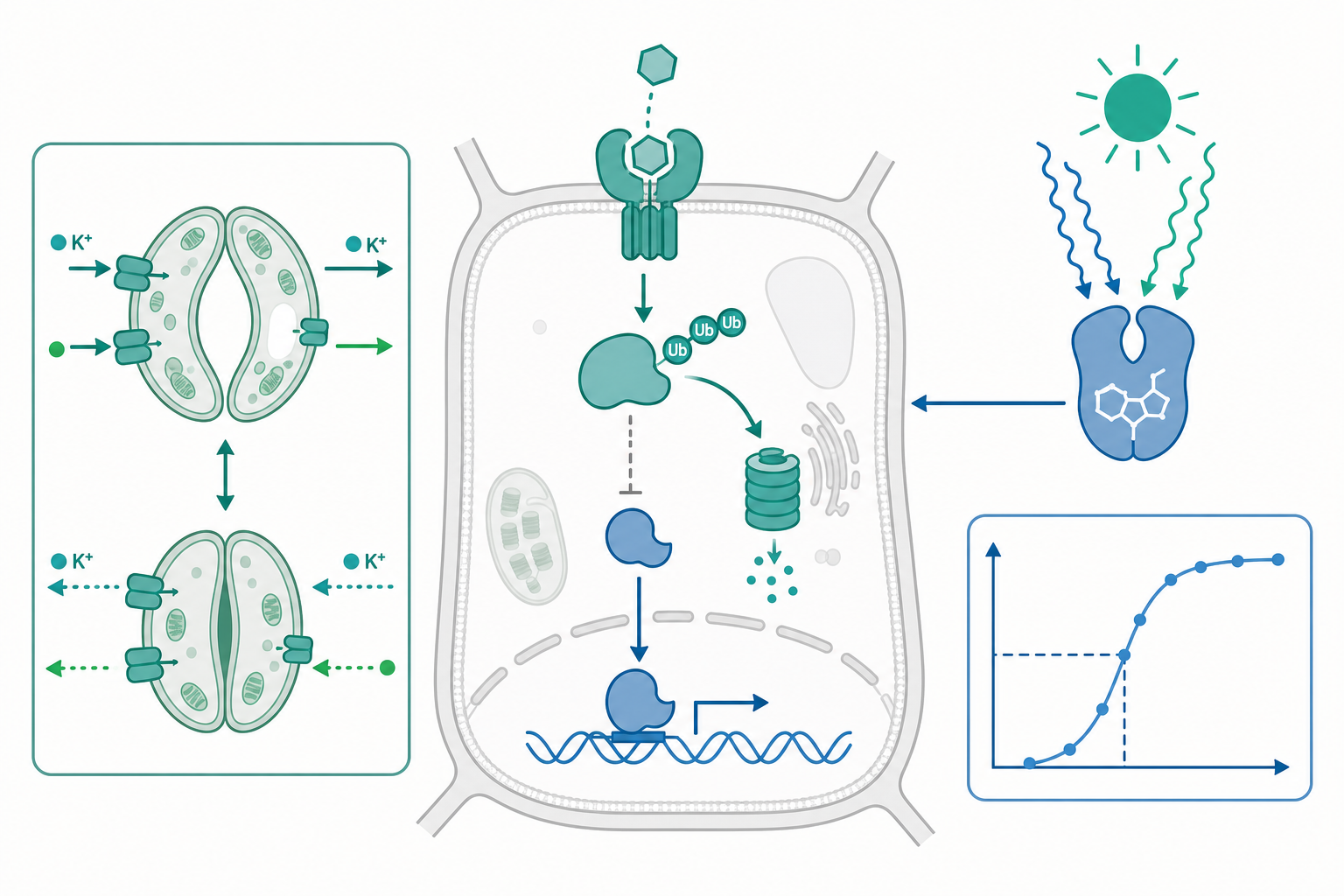
降解速率決定反應時間:一個定量小範例
「降解即訊號」帶來一個漂亮的推論:反應速度由抑制蛋白的降解速率常數 $k$ 決定。設抑制蛋白濃度 $[R]$ 在激素觸發後依一級動力學衰減:
$$\frac{d[R]}{dt} = -k\,[R] \quad\Rightarrow\quad [R](t) = [R]_0\,e^{-kt}$$
其半衰期為 $t_{1/2} = \dfrac{\ln 2}{k}$。
定量小範例:若某 DELLA 蛋白在 GA 訊號下的半衰期實測約為 10 分鐘,則 $$k = \frac{\ln 2}{t_{1/2}} = \frac{0.693}{10\ \text{min}} \approx 0.069\ \text{min}^{-1}.$$ 要把抑制蛋白降到初始量的 10%,所需時間為 $$t = \frac{\ln(1/0.10)}{k} = \frac{2.303}{0.069} \approx 33\ \text{分鐘}.$$ 這說明為何 GA 引發的去抑制反應在數十分鐘的時間尺度上展開,而非瞬間完成——降解動力學本身就是一個「低通濾波器」,過濾掉激素濃度的瞬間雜訊。
氣孔的離子計算:ABA、鈣與膜電位
環境訊號最經典的整合節點是保衛細胞(guard cell)。乾旱時離層酸(ABA)被 PYR/PYL/RCAR 受體感知,抑制 PP2C 磷酸酶、解放 SnRK2 激酶,後者磷酸化並活化 SLAC1 陰離子通道與抑制內向 K$^+$ 通道,導致溶質外流、膨壓下降、氣孔關閉。
這裡離子的「方向」由電化學梯度決定,可用能斯特方程式(Nernst equation)估算單一離子的平衡電位:
$$E_{\text{ion}} = \frac{RT}{zF}\ln\frac{[\text{ion}]_{\text{out}}}{[\text{ion}]_{\text{in}}}$$
在 25°C、單價陽離子($z=+1$)下化簡為 $E = 59\ \text{mV} \times \log_{10}\dfrac{[\text{ion}]_{\text{out}}}{[\text{ion}]_{\text{in}}}$。以鉀為例,若保衛細胞內 $[\text{K}^+]_{\text{in}}=100$ mM、外部 $[\text{K}^+]_{\text{out}}=10$ mM,則 $$E_{\text{K}} = 59 \times \log_{10}(0.1) = -59\ \text{mV}.$$ 當膜電位比 $E_{\text{K}}$ 更正時 K$^+$ 傾向外流,氣孔關閉得以進行。ABA 訊號中的鈣震盪(cytosolic Ca$^{2+}$ oscillation)則作為頻率編碼的二級訊號:CDPK 與 CBL–CIPK 系統會「讀取」鈣脈衝的頻率與振幅,決定下游通道的開閉,這是一種把連續環境變數轉成數位化指令的策略。
光與生物時鐘:訊號的時間維度
環境訊號不只有強度,還有時間結構。光敏素(phytochrome)在紅光下轉為活化型 Pfr、進入細胞核調控 PIF 轉錄因子的降解;隱花素(cryptochrome)感知藍光。這些光受體與中央生物時鐘(由 CCA1/LHY、TOC1 等構成的轉錄—轉譯負回饋環)耦合,使植物能預測黎明與黃昏。值得注意的是,激素訊號與時鐘是雙向交織的:生長素與激勃素的合成與敏感度本身受時鐘調控,造就下胚軸在特定時段的節律性伸長。換言之,植物的「反應」永遠是激素濃度、光品質、與內在時相三者的乘積,而非單一輸入的線性函數。
從交叉對話到反應網路
真正讓植物訊號難以化約的,是激素之間密集的交叉對話(crosstalk):生長素誘導乙烯合成、乙烯回頭調節生長素運輸;ABA 與 GA 在種子休眠—萌發決策上彼此拮抗,形成一個雙穩態開關。這種架構讓植物能對連續的環境輸入產生離散、近乎「全有全無」的命運決定(如萌發與否),其數學本質是含正回饋與互相抑制的非線性系統,可呈現分歧(bifurcation)與遲滯(hysteresis)。理解這些機制,意味著我們不再把激素視為單一作用的化學物質,而是當成一張運算網路的節點。
深入探討(研究所視角)
在結構生物學層次,「激素作為分子膠水」的範式已被多個高解析度結構驗證並推進。TIR1–ASK1 與 IAA、Aux/IAA degron 胜肽的共結晶結構(Tan 等人的工作)揭示 IAA 並不誘導大幅構形變化,而是單純填補受體與 degron 之間的疏水空腔、增加結合面互補性——這把「配體誘導鄰近性(ligand-induced proximity)」確立為植物受體的核心邏輯,與後來人工合成的分子膠水降解劑(molecular glue degrader)概念遙相呼應。GID1–GA–DELLA 與 COI1–JA-Ile–JAZ 的結構同樣支持此一統一框架。ABA 受體 PYL 的結構則展示「gate–latch」機制:配體結合後關閉的環狀構形,正是抑制 PP2C 活性位點的關鍵。
在系統生物學層次,研究者已能用常微分方程式網路重建氣孔 ABA 訊號的因果拓樸(Albert 等人的布林與動力學模型),定量預測單一節點突變對氣孔關閉機率的影響。這類模型的價值在於辨識網路冗餘與瓶頸節點:例如預測哪些通道是非冗餘的、可作為抗旱育種標靶。生長素的研究則發展出「運輸—反應」耦合的反應—擴散模型,PIN 蛋白的極性定位與生長素濃度形成正回饋,能自發產生周期性器官原基(葉序),把圖靈式自組織(Turing-type self-organization)的理論帶進發育生物學。
單細胞與空間體學(single-cell / spatial omics)正在重寫我們對訊號異質性的理解。單細胞 RNA-seq 顯示,過去以為「均一」的根尖組織其實在激素反應基因上呈現連續的細胞態譜(cell-state continuum),而非離散區塊;這對只看組織平均的傳統實驗提出修正。搭配可基因編碼的生物感測器——如生長素的 DII-VENUS(降解報告)、AuxSen,或鈣的 GCaMP——研究者能以亞細胞解析度、即時觀測單一細胞內激素佔據率與鈣脈衝的時空動態,把前述 $\theta = [L]/([L]+K_d)$ 的抽象佔據率變成可成像的活體讀數。
方法學前沿還包括光遺傳學與化學遺傳學工具:以光控降解(如 photo-sensitive degron)或正交化學誘導(如改造 TIR1 使其只認 cvxIAA「凸補配體」)在特定細胞、特定時刻精準擾動單一激素路徑,從相關性走向因果驗證。綜合來看,植物訊號研究正從「列舉零件」邁向「量化整個運算系統」——結構決定結合常數、結合常數餵入動力學模型、動力學模型嵌進空間反應—擴散場、再以單細胞解析度檢驗——這條由分子到系統的定量鏈,正是當代植物生理學最具張力的研究方向。