
從一個細胞到一個你:生殖與發育的奇妙旅程
配子、受精到胚胎成形,看新生命如何誕生與層層展開。
從一個細胞開始:生命如何「複製」與「重組」
想像你手上有一份珍貴的食譜,你希望它能傳給下一代。最直接的方法是整本影印——這就是「無性生殖」。細菌分裂、草莓長出走莖、扦插的盆栽,都是把自己完整複製一份,後代與親代基因幾乎一模一樣。優點是快速、不需要找伴侶;缺點是萬一這份食譜有缺陷,整個家族都會跟著倒楣。
「有性生殖」則像是把兩本食譜各撕一半,重新裝訂成一本全新的書。父親提供一半、母親提供一半,後代因此擁有獨一無二的基因組合。這樣的「重組」讓族群保有多樣性,遇到疾病或環境劇變時,總有一些個體能撐下去。生物願意付出「找對象」這麼高的成本來進行有性生殖,正是因為多樣性在演化上太重要了。
要把兩本書各撕一半再合併,數量上必須先「減半」,否則世代之間染色體會越疊越多。人類體細胞有 46 條染色體(23 對),如果精子和卵子都帶 46 條,受精後就變成 92 條,下一代再翻倍——顯然不行。解決辦法就是一種特殊的細胞分裂:減數分裂。
減數分裂:把 46 變成 23 的精密工序
一般體細胞進行的是有絲分裂:複製一次、分裂一次,產生兩個和母細胞一模一樣(46 條)的子細胞,用於生長與修復。
製造配子(精子、卵子)則用減數分裂:複製一次,卻分裂「兩次」,最後得到四個只有一半染色體(23 條)的細胞。我們可以把數量變化寫清楚:
$$2n = 46 \;\xrightarrow{\text{減數分裂 I}}\; n = 23 \;\xrightarrow{\text{減數分裂 II}}\; n = 23$$
其中 $2n$ 代表「二倍體」(成對的染色體),$n$ 代表「單倍體」(成單的染色體)。減數分裂第一次分裂時,成對的同源染色體被拉開,染色體數先減半;第二次分裂則像有絲分裂一樣把姊妹染色分體拉開,但數目維持不變。
減數分裂還做了兩件增加多樣性的事:
- 互換(crossing over):同源染色體在配對時交換片段,把父源與母源的基因「洗牌」。
- 獨立分配:每一對染色體要去哪一邊是隨機的。人類 23 對染色體,光是這個隨機就有 $2^{23}$(約 838 萬)種組合,再乘上兩位親代,難怪「世上沒有兩個人完全相同」(同卵雙胞胎除外)。
配子與受精:精子卵子的相遇
精子小而會游動,幾乎只帶遺傳物質和推進用的尾巴;卵子大而靜止,裡頭裝滿了供應早期發育所需的養分與胞器。一小一大、一動一靜,分工明確。
受精就是精子與卵子結合,把兩個單倍體(各 23 條)重新湊回二倍體(46 條)的瞬間:
$$n\,(\text{精子}) + n\,(\text{卵子}) \longrightarrow 2n\,(\text{受精卵})$$
受精卵(合子)一旦形成,會立刻啟動防止「多精入卵」的機制——卵子表面迅速產生變化,把其他精子擋在門外,確保染色體數目正確。這顆受精卵,就是一個全新個體的起點。
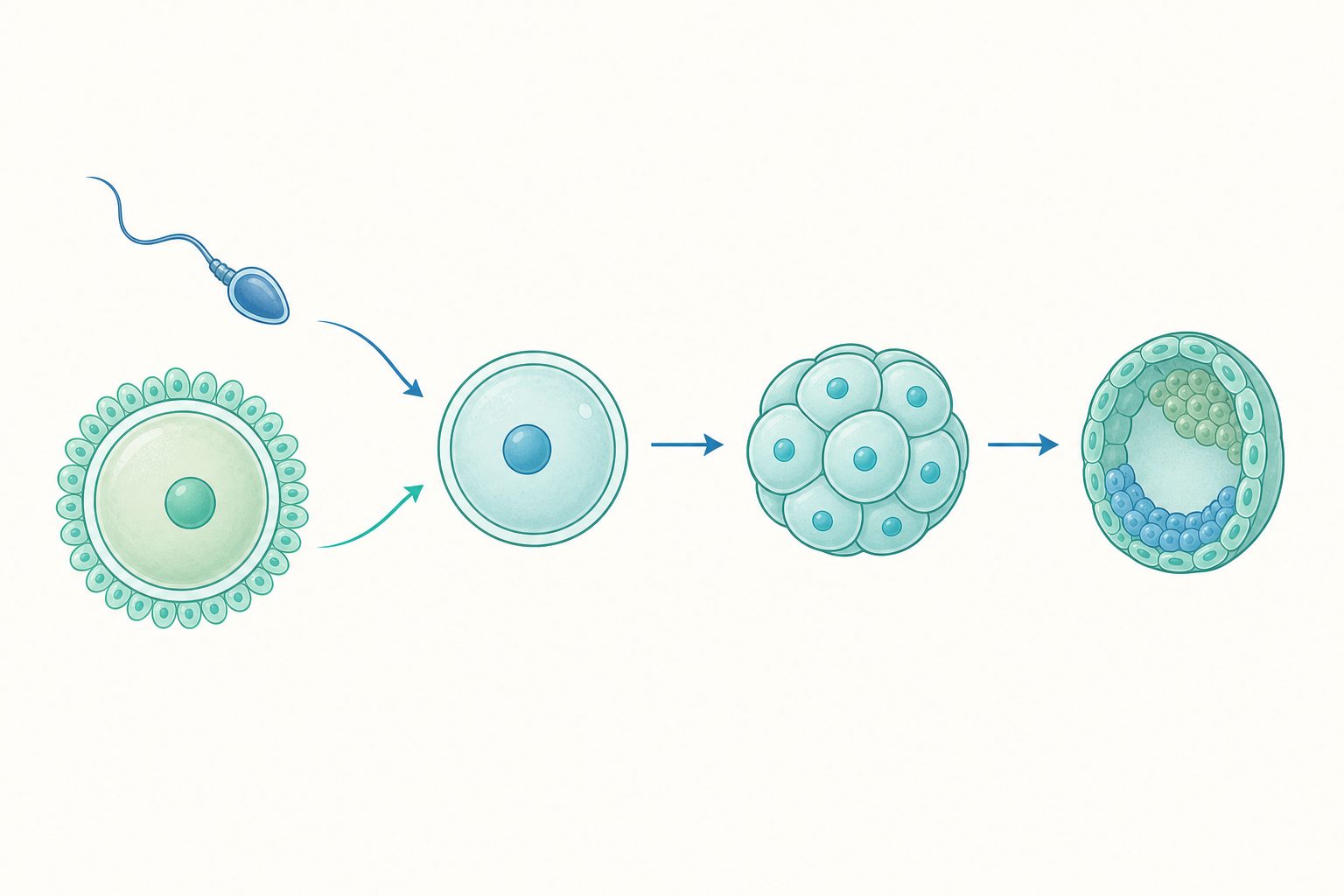
從一顆細胞到一個個體:發育的層層展開
受精卵接下來會快速分裂,這個階段稱為卵裂。有趣的是,卵裂時細胞數目增加,整體體積卻幾乎不變——就像把一塊麵團切成越來越小的小塊。
接著細胞開始「分工」。一開始的細胞具有發育成各種組織的潛能,但隨著訊號分子的引導,它們逐漸決定自己要成為皮膚、肌肉、神經還是血液——這個過程叫細胞分化。
我們可以用一組對照來理解整體流程:
| 階段 | 發生的事 | 比喻 |
|---|---|---|
| 受精 | 精+卵→受精卵(恢復 $2n$) | 兩半食譜合訂成書 |
| 卵裂 | 細胞數增加、體積不變 | 一塊麵團切小塊 |
| 分化 | 細胞各司其職 | 工人分配到不同崗位 |
| 形態發生 | 組織器官成形 | 房子蓋出格局 |
最關鍵的觀念是:身體裡幾乎每個細胞都帶有完全相同的全套基因,但神經細胞和肌肉細胞長得天差地遠。差別不在「有沒有那段基因」,而在「哪些基因被打開、哪些被關閉」。這就是發育的核心魔法——同一份食譜,不同的頁面被翻開,做出截然不同的料理。
開花與懷孕:植物與哺乳類的不同策略
不同生物把同一套原理玩出不同花樣。開花植物靠傳粉讓花粉(內含精細胞)抵達雌蕊,受精後子房發育成果實、胚珠發育成種子,種子能耐乾燥、隨風或動物傳播到遠方再萌芽。
哺乳類則選擇「體內保護」:受精卵在子宮著床,透過胎盤與母體交換養分與廢物,發育相對成熟才出生。一個把後代「打包送出」,一個把後代「貼身呵護」——沒有誰更高明,只是各自演化出適應環境的最佳解。
深入探討(研究所視角)
入門我們說「同一套基因、不同開關」,研究所層級要追問的是:開關究竟由什麼控制?答案是表觀遺傳調控(epigenetics)與形態發生素梯度(morphogen gradient)。DNA 甲基化與組蛋白修飾不改變序列,卻能穩定地開關基因,並在細胞分裂時被「記憶」傳遞,這解釋了分化後的細胞為何能維持身分。1950 年代 Conrad Waddington 提出的表觀遺傳地景(epigenetic landscape)比喻——細胞像滾下山坡的小球,逐漸落入特定山谷(命運)——至今仍是領域的核心意象。
身體軸線的建立則仰賴形態發生素:訊號分子從來源擴散形成濃度梯度,細胞依「自己所處的濃度」決定命運。經典的 法國國旗模型(French flag model)(Lewis Wolpert 提出)即描述細胞如何把連續的濃度資訊轉譯成離散的區域(藍/白/紅)。果蠅胚胎中 Bicoid 蛋白的前後梯度、脊椎動物神經管中 Sonic hedgehog 的腹背梯度,都是教科書級的實例。決定身體分節與器官位置的 Hox 基因更展現了「共線性」奇觀:基因在染色體上的排列順序,竟對應其在身體前後軸的表現位置。
胚胎發育長久以來難以追蹤「每個細胞的去向」。近年單細胞 RNA 定序(scRNA-seq)讓研究者能一次量測上萬個細胞的基因表現,重建「細胞命運軌跡(trajectory / pseudotime)」,把分化過程從黑盒子變成可繪製的分岔樹,這正是 Waddington 地景的分子版實證。CRISPR-Cas9 不僅能精準敲除特定發育基因以檢驗其功能,衍生的譜系追蹤技術(如 GESTALT)還能在基因組刻下「條碼」,事後重建細胞的家族樹。
這些工具也與其他主題深刻連結:誘導性多能幹細胞(iPSC,Shinya Yamanaka 因此獲 2012 年諾貝爾獎)證明分化並非單行道,成熟細胞可被「重新編程」回到多能狀態,直接挑戰了 Waddington 小球「只能往下滾」的直覺。發育生物學、幹細胞醫學、再生醫學與演化發育生物學(evo-devo)由此交織,共同回答一個古老的問題:一顆細胞,如何長成一個會思考的你。