
化學滲透與氧化磷酸化的分子機制
從質子驅動力到旋轉馬達:細胞呼吸末段的熱力學與結構動態
從質子的位能談起:氧化磷酸化的核心邏輯
當你呼吸時,葡萄糖最終氧化釋放的能量並不是直接「裝進」ATP,而是先被轉換成一種看似迂迴、實則精巧的形式——跨膜質子電化學梯度。1961 年 Peter Mitchell 提出的化學滲透理論(chemiosmotic theory)正是要回答這個問題:電子傳遞鏈與 ATP 合成在空間上是分離的兩件事,連接它們的是一道橫跨內膜的「質子驅動力」(proton-motive force, PMF)。粒線體不是把化學能直接傳遞,而是先把它「存」進膜兩側的離子失衡裡,再由 ATP 合成酶(ATP synthase)這台分子旋轉馬達把位能取回。
理解這套機制的關鍵,是把它當成一個熱力學與動力學耦合的系統:複合體 I–IV 把氧化還原的自由能 $\Delta G$ 轉換為跨膜電化學位能,而複合體 V 再把這份位能轉回 ATP 的磷酸鍵能。任何一環的滲漏、解偶聯或抑制,都會在量化上留下可測量的痕跡。
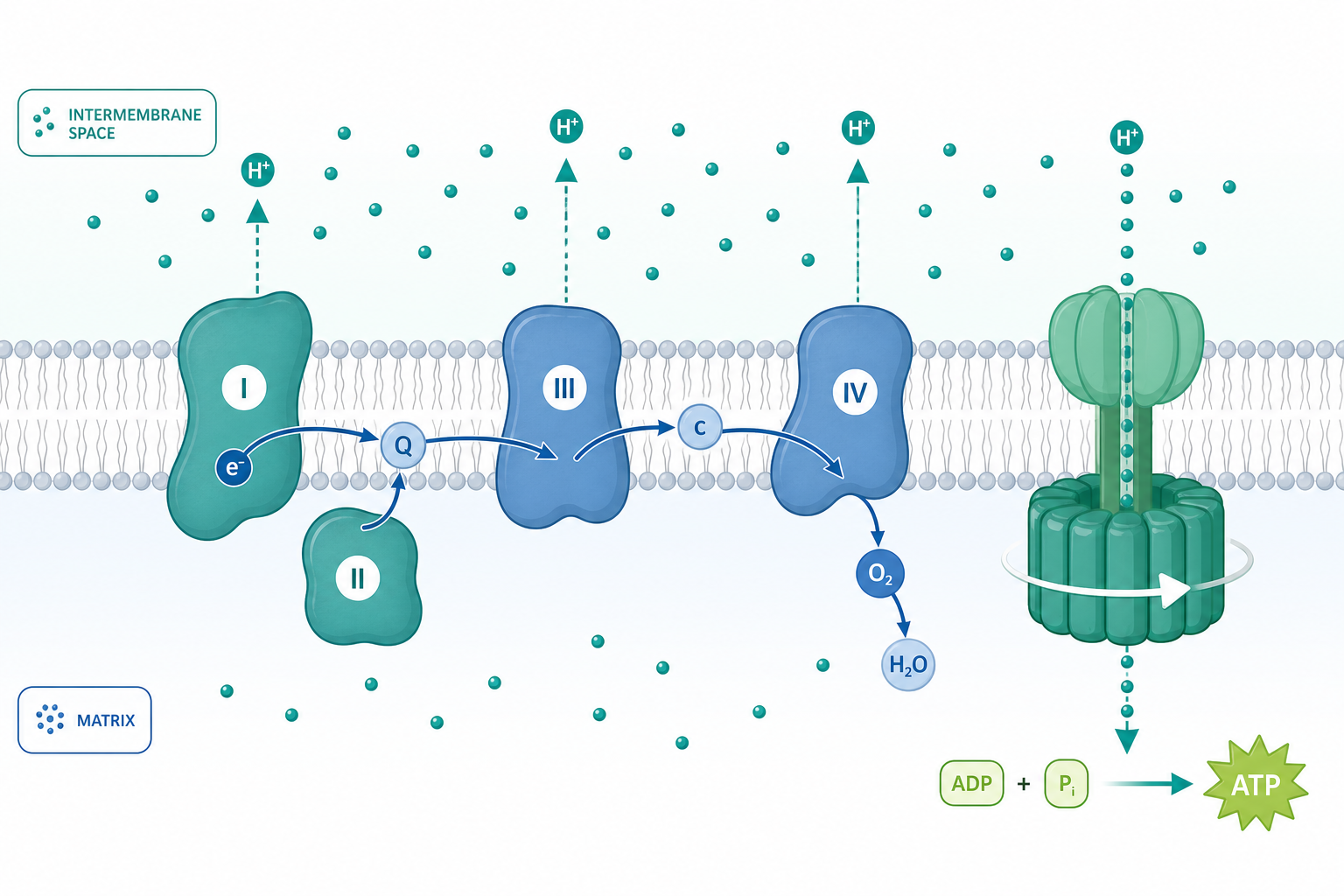
質子驅動力的定量表達
質子驅動力是化學滲透的核心物理量,由兩個分量組成:跨膜電位差 $\Delta\psi$(膜電位,內側為負)與跨膜質子濃度差 $\Delta\mathrm{pH}$。其定量形式為:
$$\Delta p = \Delta\psi - \frac{2.303\,RT}{F}\,\Delta\mathrm{pH}$$
其中 $R$ 為氣體常數、$T$ 為絕對溫度、$F$ 為法拉第常數。在 $37^\circ\mathrm{C}$($310\ \mathrm{K}$)下,$\dfrac{2.303RT}{F}\approx 61.5\ \mathrm{mV}$。典型運作中的粒線體內膜,$\Delta\psi$ 約 $150\text{–}180\ \mathrm{mV}$,$\Delta\mathrm{pH}$ 約 $0.5\text{–}1$ 單位,因此總 $\Delta p$ 約落在 $180\text{–}220\ \mathrm{mV}$。
這道梯度本質上是 Nernst 平衡概念的延伸。對單一質子物種,當系統達到平衡時,跨膜電位與濃度比由 Nernst 方程描述:
$$E = \frac{RT}{zF}\ln\frac{[\mathrm{H^+}]_{\text{out}}}{[\mathrm{H^+}]_{\text{in}}}$$
值得注意的是,$\Delta\psi$ 與 $\Delta\mathrm{pH}$ 對 ATP 合成的貢獻在熱力學上等價(都計入 $\Delta p$),但兩者的相對比例會因組織而異:植物葉綠體類囊體膜幾乎全靠 $\Delta\mathrm{pH}$,而動物粒線體則以 $\Delta\psi$ 為主。
電子傳遞鏈的幫浦化學量論
電子由 NADH 或 FADH$_2$ 進入呼吸鏈,沿著氧化還原電位梯度「下坡」流向最終受體 O$_2$。每一段下坡釋放的自由能由 Nernst 型的氧化還原電位差決定:
$$\Delta G = -nF\,\Delta E^{\circ\prime}$$
NADH/NAD$^+$ 的標準還原電位約 $-0.32\ \mathrm{V}$,O$_2$/H$_2$O 約 $+0.82\ \mathrm{V}$,總落差約 $1.14\ \mathrm{V}$。代入 $n=2$,每對電子釋放約 $-220\ \mathrm{kJ/mol}$ 的自由能。這份能量被三個幫浦複合體分段擷取:複合體 I(NADH 脫氫酶)泵出 4 個 H$^+$、複合體 III(細胞色素 bc$_1$,透過 Q 循環)泵出 4 個 H$^+$、複合體 IV(細胞色素 c 氧化酶)泵出 2 個 H$^+$。複合體 II(琥珀酸脫氫酶)不泵質子,這正是 FADH$_2$ 路徑產能較低的結構原因。
由此得到化學量論:每個 NADH 對應泵出約 10 個 H$^+$,每個 FADH$_2$ 對應約 6 個。這也解釋了為何粒線體的 P/O 比(每消耗一個氧原子合成的 ATP 數)不是過去教科書的整數 3 與 2,而是修正後的 $\approx 2.5$ 與 $\approx 1.5$——因為 ATP 合成酶的化學量論本身不是整數。
ATP 合成酶:分子旋轉馬達
ATP 合成酶(F$_o$F$_1$-ATPase)是已知最小的旋轉分子馬達,分為嵌膜的 F$_o$ 部分與伸入基質的 F$_1$ 部分。質子順著梯度通過 F$_o$ 的 c 環,驅動 c 環旋轉;中央 $\gamma$ 軸隨之轉動,並在 F$_1$ 的三個 $\beta$ 次單元上依序誘導構型變化。Paul Boyer 提出的「結合變化機制」(binding change mechanism)指出,三個催化位點循環經歷 open、loose、tight 三態:tight 態自發合成 ATP,而質子驅動的旋轉真正的功用是「推開」ATP 使其釋放。
化學量論的非整數性源於此:人類粒線體 c 環含 8 個 c 次單元,故每旋轉一圈(合成 3 個 ATP)需通過 8 個 H$^+$,即每個 ATP 需 $8/3\approx 2.67$ 個 H$^+$。再加上 ATP/ADP 與 Pi 透過載體進出基質的 $\sim 1$ 個 H$^+$ 成本,實際每個 ATP 約耗 $\sim 4$ 個 H$^+$。
定量小範例:合成一個 ATP 在熱力學上可行嗎?
設 $\Delta p = 200\ \mathrm{mV} = 0.20\ \mathrm{V}$,每個 ATP 需通過 $n_{H^+}=4$ 個質子。質子流經膜所釋放的自由能為:
$$\Delta G_{\text{H}^+} = -n_{H^+}\,F\,\Delta p = -4 \times 96\,485\ \mathrm{C/mol} \times 0.20\ \mathrm{V}$$
$$\Delta G_{\text{H}^+} \approx -77\,000\ \mathrm{J/mol} = -77\ \mathrm{kJ/mol}$$
細胞內合成 ATP 的實際自由能需求(考慮細胞內 ADP、Pi、ATP 濃度,非標準狀態)約為 $\Delta G_p \approx +50\ \mathrm{kJ/mol}$。由於 $|\Delta G_{\text{H}^+}| = 77 > 50 = \Delta G_p$,反應在熱力學上可行,且留有約 $27\ \mathrm{kJ/mol}$ 的裕度——這份裕度正是維持高速合成速率所需的「驅動過剩」。若 $\Delta p$ 因解偶聯而降到 $130\ \mathrm{mV}$,則 $|\Delta G_{\text{H}^+}|\approx 50\ \mathrm{kJ/mol}$,恰好接近平衡,ATP 合成幾乎停滯。
呼吸控制與解偶聯
在完整粒線體中,氧氣消耗速率受 ADP 供應調控,此即「呼吸控制」(respiratory control)。當 ADP 充足(state 3)時,ATP 合成酶讓質子回流,$\Delta p$ 略降,電子傳遞加速;當 ADP 耗盡(state 4)時,$\Delta p$ 升高至接近電子幫浦的「背壓」上限,呼吸減慢。這是一個自我調節的負回饋:能量需求直接透過 $\Delta p$ 與 ADP 濃度回傳到電子傳遞鏈。
解偶聯蛋白(uncoupling protein, UCP1)提供了一條繞過 ATP 合成酶的質子回流捷徑。在棕色脂肪組織中,UCP1 讓質子直接漏回基質,PMF 的位能不轉成 ATP 而是散逸為熱——這就是非顫抖性產熱(non-shivering thermogenesis)的分子基礎。化學解偶聯劑(如 2,4-DNP)則作為質子載體穿梭膜兩側,同樣崩解 $\Delta p$,使氧化與磷酸化「脫鉤」。
深入探討(研究所視角)
當代研究已將氧化磷酸化從「四個複合體加一個合成酶」的線性圖像,推進到結構動態與系統層級的整合。呼吸超複合體(respiratory supercomplexes, RCS)的發現是其中關鍵。低溫電子顯微鏡(cryo-EM)解出的「呼吸體」(respirasome, I+III$_2$+IV)顯示複合體並非自由漂浮,而是組裝成定義明確的化學量論超結構。這一安排被認為可降低活性氧(ROS)生成、限縮泛醌與細胞色素 c 的擴散路徑而提升電子傳遞效率(substrate channeling 假說),不過「固態模型」與「流體模型」之爭至今未定——同位素稀釋與單分子追蹤實驗給出的有效擴散係數,仍對超複合體是否真正限制受質流動提出質疑。
在 ATP 合成酶結構生物學方面,cryo-EM 已解析出 F$_o$F$_1$ 在不同旋轉狀態下的構型快照,揭示 c 環與 a 次單元界面上兩條「半通道」(half-channels)如何讓質子先進入、與 c 環上保守的天門冬胺酸殘基結合、隨旋轉移動近一整圈後再從另一半通道釋出。更引人注目的是,ATP 合成酶二聚體沿著嵴膜邊緣呈帶狀排列,其固有的彎折角度被認為主動塑造了粒線體嵴(cristae)的高曲率幾何,把「能量轉換」與「膜形態發生」這兩個過去分開的領域連結起來。對 滲漏電流與 ROS 的精細測量則顯示,複合體 I 的逆向電子傳遞(reverse electron transport, RET)在缺血再灌流損傷中扮演要角,琥珀酸累積會驅動電子逆流回複合體 I 而爆發 ROS,這一機制已成為心肌保護策略的標的。
系統層級上,單細胞與單粒線體技術正在改寫「均質粒線體群體」的舊假設。膜電位染劑搭配活細胞影像顯示,同一細胞內不同粒線體的 $\Delta\psi$ 存在顯著異質性,且會以「閃爍」(flickering)形式波動;這種異質性與粒線體網絡的融合—分裂動態耦合,可能是品質管控(透過 PINK1/Parkin 介導的選擇性自噬清除低 $\Delta\psi$ 的粒線體)的物理基礎。在數學建模端,源自 Magnus–Keizer 與後續整合的粒線體能量學動力學模型,把電子傳遞、PMF、ATP 合成、鈣循環與膜電位耦合成一組常微分方程,能重現呼吸控制與鈣誘發的振盪行為,並被用於預測代謝彈性與疾病狀態下的崩潰閾值。最後,結構與體學的交會——把 cryo-EM 的原子模型、蛋白質體學量化的複合體計量、與代謝體學測得的通量整合進約束式代謝模型(如 flux balance analysis 的擴充版本)——正逐步把氧化磷酸化從一條孤立通路,還原為鑲嵌在整個細胞能量網絡中、可被定量預測的節點。