
動作電位與突觸傳遞的離子機制
從能斯特平衡電位、Hodgkin–Huxley 模型到 Ca²⁺ 觸發的量子化突觸釋放
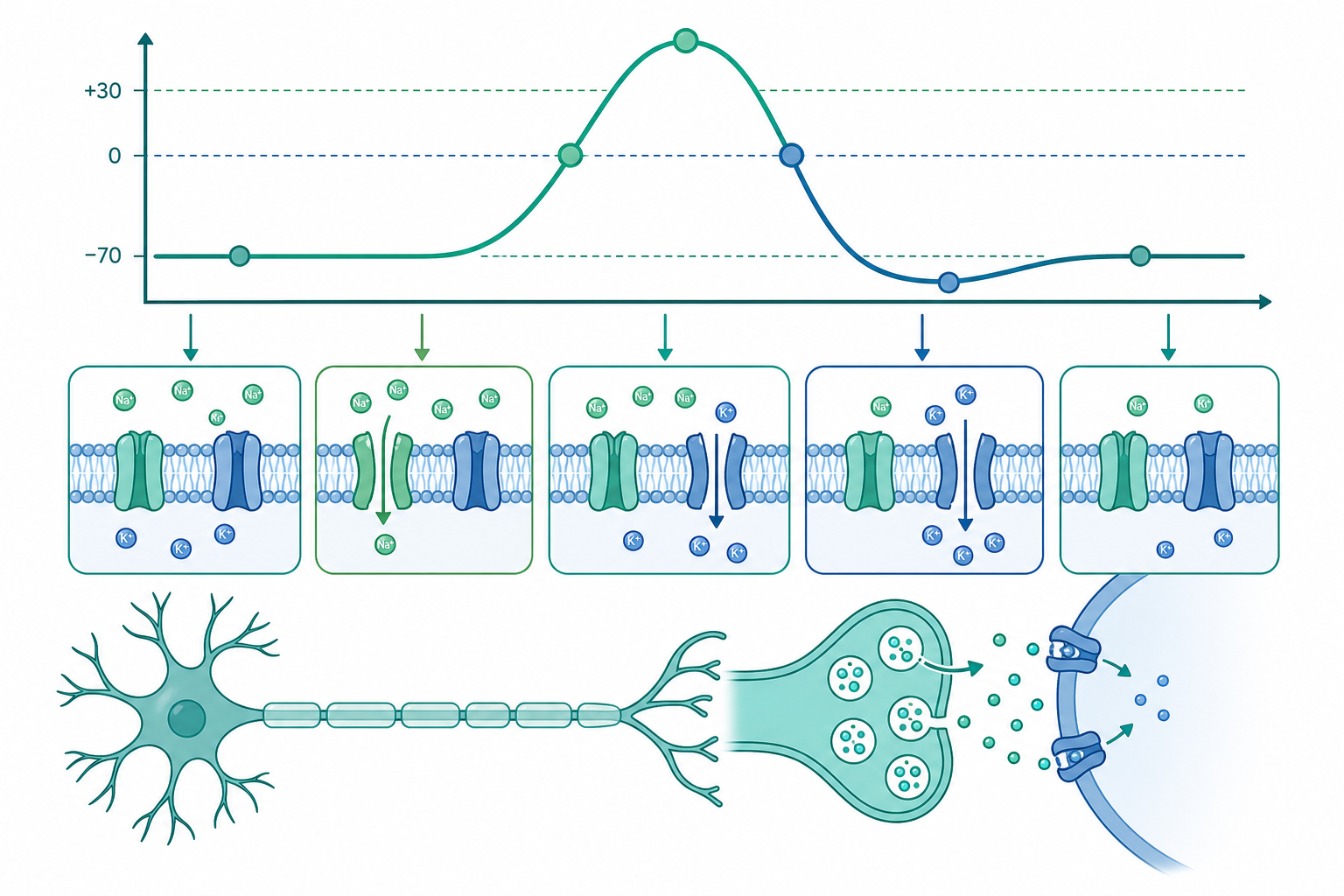
從膜電位到「全有全無」:動作電位的離子賬本
神經元為什麼能放電?最直覺的答案是:細胞膜兩側的離子分布不對稱,而膜對不同離子的通透性又會隨電壓動態改變。把這兩件事擺在一起,動作電位就不是神祕的「電火花」,而是一筆可以逐項記帳的離子收支。
靜止狀態下,神經元內部相對於外部約為 $-70\,\text{mV}$。維持這個負電位的核心是鈉鉀幫浦(Na⁺/K⁺-ATPase),它每水解一個 ATP,就把 3 個 Na⁺ 打出、2 個 K⁺ 打進,建立起跨膜的 Na⁺ 與 K⁺ 濃度梯度。典型哺乳類神經元:細胞外 $[\text{Na}^+]_o \approx 145\,\text{mM}$、$[\text{Na}^+]_i \approx 12\,\text{mM}$;$[\text{K}^+]_o \approx 4\,\text{mM}$、$[\text{K}^+]_i \approx 140\,\text{mM}$。
每一種離子都有自己的「平衡電位」,由能斯特方程(Nernst equation)給出:
$$E_{ion} = \frac{RT}{zF}\ln\frac{[ion]_o}{[ion]_i}$$
在 $37\,^{\circ}\text{C}$($T = 310\,\text{K}$)、單價陽離子($z=+1$)下,前置係數 $RT/F$ 約為 $26.7\,\text{mV}$,換成常用對數約 $61.5\,\text{mV}$。代入上述濃度:
$$E_{Na} = 61.5\,\log_{10}\frac{145}{12} \approx +66\,\text{mV},\qquad E_{K} = 61.5\,\log_{10}\frac{4}{140} \approx -95\,\text{mV}$$
靜止膜電位之所以接近 $E_K$ 而非 $E_{Na}$,是因為靜止時膜對 K⁺ 的通透性遠高於對 Na⁺ 的通透性。實際的靜止電位由 Goldman–Hodgkin–Katz(GHK)方程加權各離子的通透性求得,K⁺ 的權重最大,因此 $V_m$ 被「拉」向 $E_K$。
Hodgkin–Huxley:把通透性寫成電壓與時間的函數
1952 年 Hodgkin 與 Huxley 在槍烏賊巨軸突上完成的工作,至今仍是計算神經科學的基石。他們用電壓鉗(voltage clamp)把膜電位固定在不同值,分離出隨電壓變化的 Na⁺ 與 K⁺ 電流,並提出膜電流方程:
$$C_m\frac{dV}{dt} = -\bar{g}_{Na}\,m^3 h\,(V - E_{Na}) - \bar{g}_{K}\,n^4\,(V - E_{K}) - g_L(V - E_L) + I_{ext}$$
這裡 $m$、$h$、$n$ 是介於 0 與 1 之間的「閘門變數」:$m^3 h$ 描述 Na⁺ 通道的活化(activation,三個 m 閘)與去活化(inactivation,一個 h 閘),$n^4$ 描述 K⁺ 通道的延遲活化。每個閘門變數遵守一階動力學 $\dot{x} = \alpha_x(V)(1-x) - \beta_x(V)x$,速率常數對電壓敏感。
這組方程精準預言了動作電位的時序:
- 去極化上衝:刺激使 $V_m$ 越過閾值(約 $-55\,\text{mV}$),電壓門控 Na⁺ 通道快速活化($m$ 上升),Na⁺ 內流推 $V_m$ 衝向 $E_{Na}$,形成正回饋。
- 峰值與再極化:數毫秒內 Na⁺ 通道的失活閘($h$)關閉,同時較慢的 K⁺ 通道活化($n$ 上升),K⁺ 外流把 $V_m$ 拉回。
- 過極化與不反應期:K⁺ 通道延遲關閉造成短暫的 afterhyperpolarization;Na⁺ 通道從失活恢復前,存在絕對與相對不反應期,這正是動作電位單向傳導、不會回頭的分子基礎。
值得注意的是「全有全無」並非每次流入等量電荷,而是只要超過閾值,正回饋就會把反應推到飽和峰值;閾值以下則無法觸發。資訊因此編碼在頻率而非幅度。
跳躍式傳導與一筆電荷小計
在有髓鞘的軸突上,動作電位只在 Ranvier 節(node of Ranvier)重新生成,於節間以電纜方式被動傳播,形成跳躍式傳導(saltatory conduction),大幅提升速度並節省離子泵的能量開銷。
來算個定量小範例:一段膜電容 $C_m \approx 1\,\mu\text{F/cm}^2$,動作電位幅度約 $\Delta V = 100\,\text{mV} = 0.1\,\text{V}$。單位面積去極化所需電荷:
$$Q = C_m\,\Delta V = (1\times 10^{-6}\,\text{F/cm}^2)(0.1\,\text{V}) = 1\times 10^{-7}\,\text{C/cm}^2$$
換算成 Na⁺ 離子數($F = 96485\,\text{C/mol}$、$N_A = 6.02\times10^{23}$):
$$\frac{Q}{F}\cdot N_A = \frac{10^{-7}}{96485}\times 6.02\times10^{23} \approx 6\times10^{11}\ \text{ions/cm}^2$$
這個數字看似龐大,但相對於胞內 Na⁺ 總量只是極小擾動——單次放電幾乎不改變整體濃度,所以才能連續放電上千次而不立刻耗盡梯度。長期則由鈉鉀幫浦持續償還這筆「電荷貸款」。
突觸傳遞:把電訊號譯成化學再譯回電
動作電位抵達軸突末梢後,必須跨越突觸間隙。化學突觸的關鍵是把膜電位變化轉換成 Ca²⁺ 訊號:
- 去極化開啟電壓門控 Ca²⁺ 通道,$[\text{Ca}^{2+}]_o \approx 2\,\text{mM}$ 而 $[\text{Ca}^{2+}]_i$ 僅約 $100\,\text{nM}$,巨大梯度驅動 Ca²⁺ 瞬間內流。
- Ca²⁺ 結合於 synaptotagmin,觸發 SNARE 複合體(syntaxin、SNAP-25、synaptobrevin)介導的突觸囊泡融合,神經傳遞物以量子化(quantal)方式釋放。Katz 等人證明釋放是離散的「量子」單位,其平均數遵守泊松統計。
- 神經傳遞物(如麩胺酸、GABA)擴散結合突觸後受體。興奮性受體(如 AMPA、NMDA)開啟陽離子通道,產生興奮性突觸後電位(EPSP);抑制性受體(GABA_A)開啟 Cl⁻ 通道,因 $E_{Cl}$ 接近靜止電位而抑制放電。
突觸後膜把眾多 EPSP 與 IPSP 在時間與空間上整合,一旦軸丘的去極化越過閾值,就再次點燃全有全無的動作電位。NMDA 受體尤其關鍵:它同時是配體門控與電壓門控(靜止時被 Mg²⁺ 堵塞),需「突觸前釋放麩胺酸」與「突觸後去極化」同時滿足才導通 Ca²⁺,因而成為偵測前後活動巧合的分子裝置,是長期增益(LTP)等突觸可塑性的核心。
深入探討(研究所視角)
主體把通道當成具有平滑機率的「閘門變數」,但結構生物學已把這些變數還原成蛋白構象。電壓門控鈉通道(Naᵥ)與鉀通道(Kᵥ)屬於同一超家族,核心是四個含 S1–S6 跨膜片段的結構域;S4 螺旋帶有規律排列的正電荷殘基,構成「電壓感測器」。MacKinnon 實驗室以 X 光晶體學解出 KcsA 與 Kᵥ 通道,揭示選擇性過濾器由主鏈羰基氧模擬水合殼層,使 K⁺(而非較小的 Na⁺)以近擴散極限的速率通過——這項對離子通道選擇性與門控機制的結構解釋獲 2003 年諾貝爾化學獎。近年冷凍電顯(cryo-EM)更直接捕捉到 Naᵥ 與 Caᵥ 在失活、開放等狀態的原子構象,把 Hodgkin–Huxley 的 $m$、$h$、$n$ 對應到具體的螺旋滑移與失活閘(如 Naᵥ 的 IFM motif 像「鉸鏈蓋」塞住孔道)。
在訊號傳遞的下游,突觸釋放機制也被推進到結構與單分子層次。Südhof 等人闡明 Ca²⁺ 感測、SNARE 拉鏈式組裝與 complexin 對融合的鉗制/觸發調控(2013 年諾貝爾生理醫學獎)。全內反射螢光(TIRF)與光遺傳學讓研究者得以在單一囊泡尺度觀測融合動力學,把 Katz 的量子假說落實到可視化的單囊泡事件。
系統與計算層面,HH 形式被推廣為多隔室(multi-compartment)電纜模型,結合形態重建的樹突,模擬樹突棘上的非線性整合與 NMDA 尖峰(NMDA spike);NEURON 等模擬平台已能在形態真實的神經元上重現主動樹突計算。隨機性也被正視:單通道層次的開關是離散事件,Markov 鏈模型取代確定性 HH,可解釋通道雜訊如何影響閾值附近的放電可靠度。
最前沿的是「橋接尺度」的整合。Patch-seq 把單細胞膜片鉗的電生理參數、形態與單細胞 RNA 定序(轉錄體)對齊,使 Allen Institute 等團隊得以用離子通道基因(如 SCN、KCN、CACNA 家族)的表現量,解釋不同神經元類型為何具有不同的放電特徵與閾值。離子通道病(channelopathies)研究則反向印證機制:Naᵥ1.1(SCN1A)功能喪失與 Dravet 癲癇相關、Caᵥ 突變與偏頭痛及小腦失調相連,把單一胺基酸的構象改變一路推到行為層級的表型。這些工作共同把「離子賬本」從槍烏賊軸突,延伸成可量化、可定序、可建模的多體學圖譜。