
演化發育生物學:從調控網路到形態多樣性的量化解析
為何形態演化主要來自既有基因「使用方式」的改變,而非新基因的出現
從「基因的差異」到「身體的差異」
為什麼黑猩猩與人類有約 98.8% 的基因組相同,外形與行為卻天差地別?為什麼蝙蝠的翼、鯨魚的鰭、人類的手,骨骼排列卻同源?演化發育生物學(Evolutionary Developmental Biology,簡稱 Evo-Devo)的核心洞見是:形態的演化,大多不是來自「新基因的出現」,而是來自「既有基因在發育時程中被使用方式的改變」。換言之,演化作用的對象往往不是蛋白質序列本身,而是控制「何時、何地、用多少」的調控邏輯。本文不停留在這個直覺,而是進入分子機制、量化模型與最前沿的觀測層次。
Evo-Devo 整合了三條傳統上分離的線索:發育生物學(個體如何從受精卵建構出來)、演化生物學(族群中遺傳變異如何被篩選)、以及比較基因體學(不同物種的基因組如何分歧)。它要回答的不是「哪個基因突變了」,而是「發育的調控網路如何被微調,使得族群層級的變異得以放大為跨物種的形態鴻溝」。
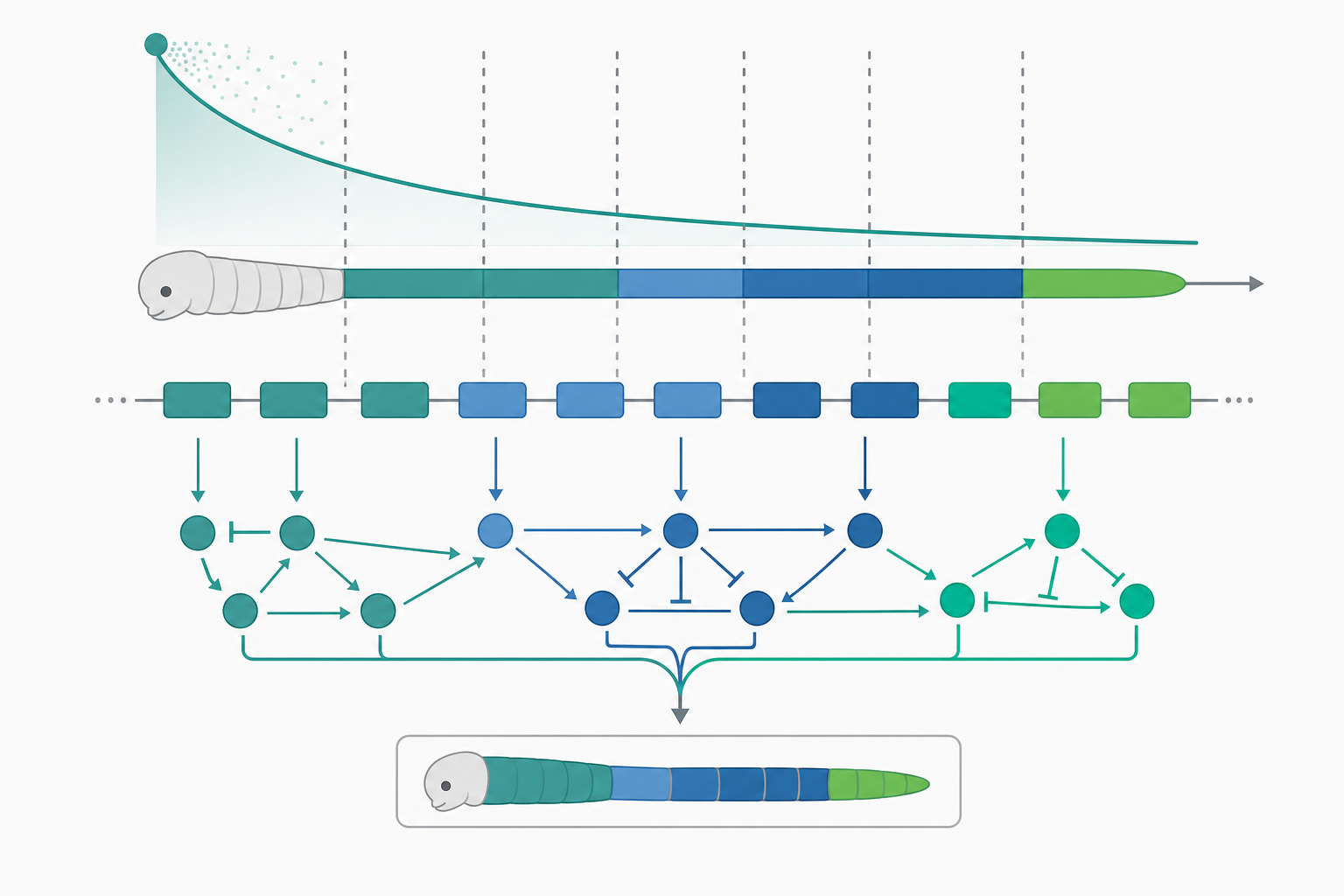
Hox 基因與發育工具箱的深層同源
Evo-Devo 的奠基性發現之一,是動物界共用一套高度保守的「發育工具箱基因」(developmental toolkit genes),其中最著名的是 Hox 基因家族。Hox 基因編碼含有同源異形域(homeodomain)的轉錄因子,沿著前後軸(anterior-posterior axis)以共線性(colinearity)方式排列——基因在染色體上的物理順序,對應其在身體軸上的表現位置與時序。果蠅、小鼠到人類都共享此邏輯,這就是「深層同源」(deep homology)。
關鍵在於:Hox 基因的蛋白質序列極度保守,物種間的形態差異主要來自其表現域邊界的位移。例如蛇類軀幹的肋骨化、昆蟲附肢數目的改變,往往對應 Hox 表現域(如 Ubx)在體節上的擴張或收縮,而非 Hox 蛋白本身的結構突變。這支持了 Evo-Devo 的中心命題:順式調控(cis-regulatory)演化是形態多樣性的主要引擎。順式調控元件(如增強子 enhancer)的突變具有模組化、低多效性(pleiotropy)的優勢——改一個增強子只影響特定組織特定時期的表現,不會像編碼區突變那樣牽動該蛋白所有功能,因此較不易被淨化選擇剔除。
形態建構的定量基礎:形態素梯度與閾值
發育中的位置資訊(positional information)常由形態素(morphogen)的濃度梯度編碼。經典如 Bicoid 在果蠅前後軸形成指數遞減梯度,下游基因依「閾值」被開或關,把連續的濃度差轉譯為離散的體節邊界。這正是 Lewis Wolpert 的「法國國旗模型」。
形態素梯度可用反應—擴散框架近似。最簡的穩態擴散—降解模型中,濃度 $C(x)$ 沿位置 $x$ 滿足
$$D\frac{d^2 C}{dx^2} - k\,C = 0$$
其中 $D$ 為擴散係數、$k$ 為降解速率常數。其解為指數衰減 $C(x) = C_0\, e^{-x/\lambda}$,特徵長度 $\lambda = \sqrt{D/k}$ 決定了梯度的「空間尺度」。Evo-Devo 的洞見是:$\lambda$ 的微小演化變化(例如降解酶活性改變使 $k$ 變動),就能整體縮放或拉伸身體圖式,提供一個無須新增基因即可改變比例的旋鈕。
下游基因對形態素的反應常呈現協同性(cooperativity),可用 Hill 方程描述被啟動的比例 $\theta$:
$$\theta = \frac{C^n}{K^n + C^n}$$
$n$ 為 Hill 係數,反映轉錄因子結合的協同程度;$K$ 為半飽和濃度。$n$ 越大,反應曲線越陡,閾值越「開關化」,邊界越銳利。演化可透過調整增強子上轉錄因子結合位點的數目與親和力來改變 $n$ 與 $K$,從而銳化或模糊形態邊界。
定量小範例:閾值位置如何隨梯度尺度移動
假設某下游基因在形態素濃度達到啟動閾值 $C_T = 0.1\,C_0$ 時才開啟邊界。源頭濃度 $C_0$ 固定,初始特徵長度 $\lambda_1 = 50\ \mu\mathrm{m}$。邊界位置 $x_T$ 滿足 $C_T = C_0 e^{-x_T/\lambda}$,即
$$x_T = -\lambda \ln\!\left(\frac{C_T}{C_0}\right) = -\lambda \ln(0.1) = \lambda \cdot 2.303$$
故 $x_{T,1} = 50 \times 2.303 \approx 115\ \mu\mathrm{m}$。
今一個演化突變使降解速率減半,$k \to k/2$,則 $\lambda = \sqrt{D/k}$ 增為 $\lambda_2 = 50 \times \sqrt{2} \approx 70.7\ \mu\mathrm{m}$。新邊界位置
$$x_{T,2} = 70.7 \times 2.303 \approx 163\ \mu\mathrm{m}$$
邊界外移約 48 微米——僅僅改變一個降解參數,就把這個結構的相對位置整體後推。若同一梯度控制多個閾值基因,整個器官原基會等比例放大。這示範了 Evo-Devo 的量化精神:單一調控參數的連續變動,如何映射為形態的離散重組。
族群層級:調控變異如何被固定
Evo-Devo 不只是發育機制,也必須接回族群遺傳學。一個改變增強子的突變要造成跨物種形態差異,必須先在族群中達到可觀頻率。在無選擇、隨機交配的理想族群中,等位基因頻率遵循 Hardy–Weinberg 平衡:對頻率為 $p$ 與 $q=1-p$ 的兩個等位基因,基因型頻率為
$$p^2 + 2pq + q^2 = 1$$
真實的調控突變則受方向性選擇與遺傳漂變影響。Evo-Devo 強調順式調控突變相對於編碼突變的選擇優勢:因其組織特異性,適應度代價(fitness cost)較低,等位基因可在不嚴重損害其他功能下被正向選擇推升。著名的實證是棘背魚(threespine stickleback)腹鰭棘刺退化——同一基因 Pitx1 的組織特異性增強子反覆獨立缺失,導致淡水族群骨盆結構消失,而 Pitx1 編碼區保持完整(編碼區突變會致命,因該基因亦參與其他器官發育)。這是「平行演化由相同調控位點反覆命中」的經典案例。
異時性、異位性與形態空間
形態演化可拆解為幾個可量化的維度:異時性(heterochrony,發育事件時序改變)、異位性(heterotopy,表現空間位置改變)、異速生長(allometry,部位相對生長速率改變)。異速生長常以對數線性關係描述:
$$\log y = \log a + b \log x$$
其中 $x$ 為體型、$y$ 為某器官大小、$b$ 為異速指數。$b=1$ 為等速;$b>1$ 表示該器官隨體型放大而超比例增大(如雄鹿角)。演化只需改變 $b$,就能在不發明新結構的情況下,沿既有的生長軌跡探索形態空間。Evo-Devo 由此把「形態如何可能改變」(developmental possibility)與「形態實際改變了多少」(evolutionary realization)連成一條可測量的鏈。
深入探討(研究所視角)
當代 Evo-Devo 已從「比較表現域」推進到機制解析與全基因組尺度的定量,幾條前沿值得研究生關注。
順式調控的結構與生化解碼。 增強子並非被動的結合位點清單,而是具有特定排列文法(grammar)的調控邏輯。轉錄因子如何在開放染色質中協同佔據增強子,可用熱力學結合模型描述:佔據機率取決於結合自由能 $\Delta G$,由 $K_d = e^{\Delta G / RT}$ 與位點濃度共同決定,多位點間的協同(如 pioneer factor 先開染色質)改變了有效 $\Delta G$。冷凍電子顯微術(cryo-EM)近年解析了轉錄因子—核小體—中介複合體(Mediator)的結構,揭示「同源異形域如何辨識 DNA 形狀(DNA shape readout)而非僅辨識序列」,這為「序列幾乎不變卻造成表現量改變」提供了結構基礎。
單細胞與時空體學重寫發育圖譜。 單細胞 RNA 定序(scRNA-seq)、單細胞 ATAC-seq 與空間轉錄體(spatial transcriptomics)讓研究者得以重建細胞命運軌跡(trajectory)與分支點的調控狀態,而非僅看整體組織平均。跨物種比較的單細胞圖譜(如比較人腦與其他靈長類皮質發育)正在定位「哪些細胞類型、在哪個發育窗口、哪些增強子被差異活化」,把 Evo-Devo 的解析度推到細胞型別層級。RNA 速度(RNA velocity)與譜系追蹤(lineage tracing,如 CRISPR 分子條碼)進一步把「狀態」轉為「動力學」。
基因調控網路的系統生物學模型。 把發育視為基因調控網路(GRN)的動力系統,是 Eric Davidson 等人奠定的範式。網路狀態可用常微分方程組描述,每個調控節點的表現 $g_i$ 隨時間變化:
$$\frac{dg_i}{dt} = \beta_i\, H(\,\{g_j\}\,) - \gamma_i\, g_i$$
其中 $\beta_i$ 為最大合成速率、$\gamma_i$ 為降解速率、$H$ 為整合上游輸入的調控函數(常用 Hill 型)。網路中的「核心調控迴路(kernels)」高度保守、改動即致命,解釋了體軸基本圖式的穩定;而周邊的「插件(plug-ins)」與末端分化基因battery 則容許演化彈性。這種模組化拓樸是「保守與創新並存」的系統層解答。發育的穩健性(canalization)也可在此框架量化——網路對參數擾動的不敏感性,正是 Waddington 表觀遺傳地景(epigenetic landscape)的數學再陳述。
新近機制發現的方向。 包括:增強子的冗餘與穩健性(shadow enhancers 在環境擾動下緩衝表現變異);三維基因組架構(TAD、染色質環)如何限制增強子—啟動子配對,使調控突變的效應「拓樸受限」;以及非編碼 RNA 與表觀遺傳標記在跨世代調控中的角色。對研究生而言,最具張力的開放問題是:發育約束(developmental constraint)究竟是限制了演化,還是「引導」了演化方向(developmental bias)——亦即形態空間中並非所有方向等機率可及,發育系統的結構本身偏置了變異的供應,使演化在某些軸向上更容易發生。這把 Evo-Devo 從「事後解釋」推向「可預測」的科學雄心。