
族群遺傳與 Hardy–Weinberg 平衡:從虛無模型到基因組推論
用機率推導、選汰係數與合併理論,量化看不見的演化力量
為什麼一個族群的基因會「靜止」,又為什麼總是不會?
直覺上,我們以為顯性等位基因(dominant allele)會隨世代越來越普遍、隱性等位基因終將消失。但這個直覺是錯的。族群遺傳學最深刻的起點,正是一個反直覺的結論:在一組理想條件下,等位基因頻率(allele frequency)與基因型頻率(genotype frequency)會世世代代維持不變。這個「不變」就是 Hardy–Weinberg 平衡。它的真正價值不在於描述自然界(自然界幾乎沒有族群真正處於平衡),而在於它提供了一個虛無模型(null model):當實際觀測偏離它,我們就知道有演化力量正在運作,並能據此量化那股力量的方向與強度。
對基礎學習者而言,重點不再是「複習 $p^2 + 2pq + q^2 = 1$」,而是理解這條式子背後的機率推導、它依賴的五個假設如何各自對應一種演化機制,以及如何把偏離量化成選汰係數、近交係數與遷移率。
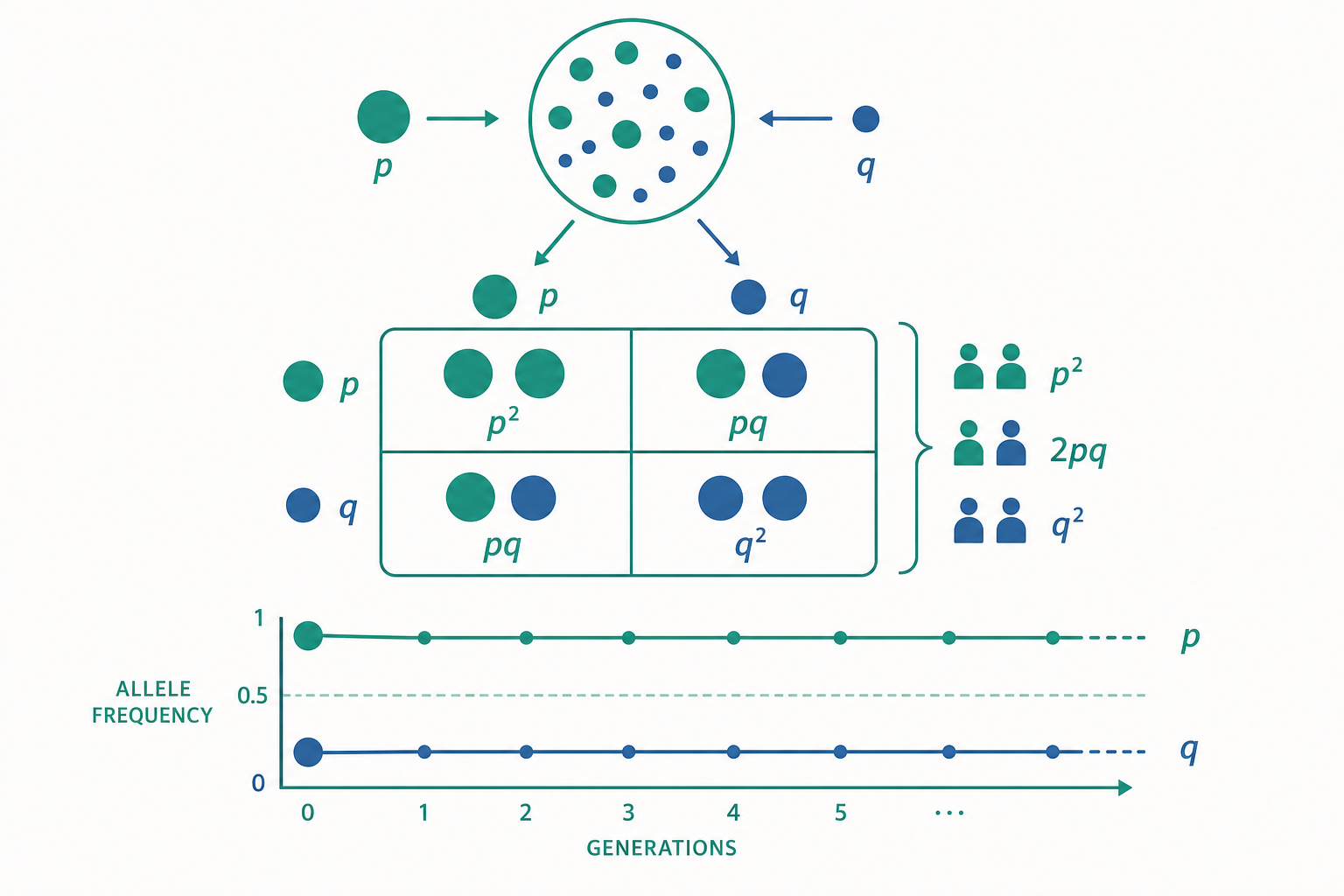
從等位基因頻率到基因型頻率:機率的嚴格推導
考慮單一基因座(locus)上的兩個等位基因 $A$ 與 $a$,頻率分別記為 $p$ 與 $q$,且 $p + q = 1$。Hardy–Weinberg 定律宣稱:在隨機交配(random mating)下,下一世代的基因型頻率為
$$ f(AA) = p^2,\quad f(Aa) = 2pq,\quad f(aa) = q^2. $$
這並非定義,而是一個機率結果。在隨機交配下,配子(gametes)的結合可視為從同一個配子池(gamete pool)獨立抽取兩枚配子。抽到 $A$ 的機率為 $p$,抽到 $a$ 為 $q$。因此抽到兩枚 $A$ 的機率是 $p \times p = p^2$;抽到一 $A$ 一 $a$ 有兩種排列($A$ 來自父或母),故為 $2pq$。這正是二項式展開:
$$ (p + q)^2 = p^2 + 2pq + q^2 = 1. $$
關鍵洞見:一個世代的隨機交配即足以使任意起始基因型頻率回到 H–W 比例,之後便維持恆定。也就是說平衡是「一步到位」的(針對單一常染色體基因座、雌雄頻率相同的情形)。這背後的數學是離散世代下的不動點(fixed point)。
成立的五個假設環環相扣:(1) 無突變、(2) 無天擇、(3) 無遷移(基因流)、(4) 無遺傳漂變(即族群無限大)、(5) 隨機交配。每個假設被打破,都對應一個可量化的演化過程。
偏離平衡:把演化力量寫成方程式
天擇(selection)。 設三種基因型 $AA$、$Aa$、$aa$ 的相對適存度(relative fitness)為 $w_{11}, w_{12}, w_{22}$。以 $A$ 為有利等位基因、$a$ 為有害且選汰係數(selection coefficient)為 $s$ 的方向選汰為例,令 $w_{11}=1, w_{12}=1, w_{22}=1-s$(顯性有利)。一世代後 $q$ 的變化為
$$ \Delta q = \frac{-s\,p\,q^2}{1 - s\,q^2}. $$
注意分子含 $q^2$:當 $a$ 已稀有,$q^2$ 極小,$\Delta q$ 趨近於零。這解釋了為何天擇極難清除隱性有害等位基因——稀有時它幾乎全藏在雜合子(heterozygote)中而不受選汰。這也是許多隱性遺傳疾病在族群中持續存在的核心原因。
突變(mutation)。 設正向突變率 $A \to a$ 為 $\mu$、回復突變率為 $\nu$,則平衡頻率為 $\hat{q} = \mu/(\mu+\nu)$。突變率通常極小($\sim 10^{-6}$ 至 $10^{-5}$/座/世代),單獨作用緩慢,但它是所有新變異的最終源頭。
遺傳漂變(genetic drift)。 在有限族群(有效族群大小 $N_e$)中,等位基因頻率每世代隨機抽樣而波動。$p$ 的抽樣變異數為
$$ \mathrm{Var}(\Delta p) = \frac{p\,q}{2N_e}. $$
族群越小,波動越大,越易導致等位基因隨機固定(fixation)或消失。漂變是中性演化(neutral theory)的引擎。
近交(inbreeding)。 非隨機交配不改變等位基因頻率,但改變基因型分布。引入近交係數 $F$:
$$ f(AA)=p^2+Fpq,\quad f(Aa)=2pq(1-F),\quad f(aa)=q^2+Fpq. $$
$F>0$ 時雜合子減少、同型合子增多,使隱性性狀更易顯現。
定量小範例:用 H–W 估計帶因者頻率
假設某常染色體隱性遺傳疾病在某族群的發病率為每 10,000 人有 1 人,即 $f(aa)=q^2=1/10000$。求帶因者(雜合子)頻率。
步驟一: 由 $q^2 = 0.0001$ 解出 $q$:
$$ q = \sqrt{0.0001} = 0.01. $$
步驟二: 由 $p + q = 1$ 得 $p = 0.99$。
步驟三: 帶因者頻率 $f(Aa)=2pq$:
$$ 2pq = 2 \times 0.99 \times 0.01 = 0.0198 \approx 1.98\%. $$
換言之,雖然發病者僅萬分之一,但約每 50 人就有 1 人是無症狀帶因者。雜合子數量遠多於患者(比值約 $2p/q \approx 198$ 倍),這正是上一節「天擇難清除隱性等位基因」的數量級註腳,也是遺傳諮詢中帶因者篩檢的理論基礎。
連鎖不平衡:當基因座不再獨立
單座 H–W 只是起點。真實基因組上多個基因座彼此相鄰,重組(recombination)未必能打散它們的組合。連鎖不平衡(linkage disequilibrium, LD) 量化兩座位等位基因非隨機共現的程度,定義為
$$ D = f(AB) - p_A\,p_B, $$
其中 $f(AB)$ 是單套體(haplotype)$AB$ 的觀測頻率,$p_A p_B$ 是若獨立應有的期望值。每世代 LD 以重組率 $r$ 衰減:$D_{t} = (1-r)^t D_0$。LD 是全基因組關聯分析(GWAS)能用標記 SNP 定位致病變異的根本前提,也承載族群歷史(瓶頸、混合、選汰掃蕩)的訊號。
深入探討(研究所視角)
當代族群遺傳學早已從單座代數走向基因組尺度的合併理論與推論統計,以下幾條前沿值得研究所階段深究。
合併理論(coalescent theory)。 Kingman 於 1982 年提出的合併過程,把視角從「前向」(forward-in-time,追蹤頻率演變)翻轉為「後向」(backward-in-time,追蹤譜系合併)。在中性、$N_e$ 恆定的 Wright–Fisher 模型下,現存 $k$ 條譜系中任兩條往回追溯到最近共同祖先的等待時間服從指數分布,期望為 $\frac{4N_e}{k(k-1)}$ 世代。合併框架的威力在於它讓我們能高效模擬序列資料、並從觀測到的核苷酸多樣性 $\pi$ 與分離位點數 $S$ 反推 $\theta = 4N_e\mu$。Tajima's $D$ 統計量正是比較這兩種 $\theta$ 估計值的差異,用以偵測偏離中性的選汰或族群大小變動:$D<0$ 暗示近期擴張或正選汰掃蕩,$D>0$ 暗示平衡選汰或族群收縮。
選汰掃蕩與背景選汰的基因組標誌。 強正選汰使有利變異快速固定,會「搭便車」(genetic hitchhiking)連帶固定鄰近中性變異,在基因組留下局部多樣性低谷與長單套體區塊。iHS、XP-EHH、FST 離群掃描等統計量即用以定位這類選汰足跡(selective sweep signatures)。人類乳糖耐受(LCT/MCM6 區域)、高海拔適應(EPAS1,藏族)皆是教科書級的近期正選汰實例,其機制已連結到特定調控變異與生理表現型。
結構化與祖源推論。 真實族群並非單一隨機交配池,而是有層次結構(population structure)。Pritchard 等人的 STRUCTURE 與後續的 ADMIXTURE、主成分分析(PCA)能從全基因組 SNP 推斷個體的祖源比例與混合歷史。這在 GWAS 中至關重要——未校正的族群分層會產生假關聯(confounding),故需以主成分或混合模型(linear mixed model)校正。
單細胞與多體學的新層次。 傳統族群遺傳處理生殖細胞系(germline)變異的世代傳遞;近年單細胞定序揭示體細胞(somatic)演化同樣遵循族群遺傳規律:腫瘤內不同細胞株如同競爭的等位基因,可用合併與選汰模型刻畫其譜系與適存度差異。將 H–W 與選汰框架延伸到體內細胞族群,是癌症演化(cancer evolution)研究的理論骨幹。同時,整合表觀基因體、轉錄體的多體學資料,使我們得以區分變異是經由蛋白序列改變(編碼區)或表現量調控(順式調控元件)影響表現型,呼應前述乳糖耐受案例中調控變異主導適應的發現。族群遺傳學的核心方程式並未過時,而是被嵌入越來越高維、越來越接近機制的推論架構之中。