
從一口三明治到一身能量:消化與吸收的奇妙旅程
食物如何被拆解、吸收,轉成身體能用的養分?
為什麼我們要吃東西?
想像你的身體是一座永不打烊的城市。城市需要兩樣東西才能運作:能量(讓燈亮著、機器轉著)和建材(修補老舊建築、蓋新房子)。食物,就是這座城市每天運進來的物資。但問題是——運進來的物資是「整箱整箱」的,沒辦法直接使用。一塊雞排、一碗白飯、一杯牛奶,這些都是由巨大的分子組成的,太大了,根本進不了我們的細胞。
於是身體做了一件很聰明的事:把大分子拆成小零件。這個「拆解」的過程,就是消化;把拆好的小零件搬進身體裡,就是吸收。營養與消化,講的其實就是這一條「從食物到細胞」的旅程。
我們吃進去的食物,主要分成三大類巨量營養素:醣類(碳水化合物)、蛋白質、脂質(脂肪)。它們各自被拆成不同的小零件:
- 醣類 → 拆成單醣(如葡萄糖 $C_6H_{12}O_6$)
- 蛋白質 → 拆成胺基酸
- 脂質 → 拆成脂肪酸與甘油
這些小零件進入細胞後,有些被「燒掉」產生能量,有些則被拿去當建材。把食物變成能量的最核心反應,就是細胞呼吸:
$$C_6H_{12}O_6 + 6O_2 \rightarrow 6CO_2 + 6H_2O + \text{能量(ATP)}$$
也就是說,你早餐吃的那碗飯,最後其實是被你「呼吸」掉的——它的碳變成二氧化碳呼出去,能量則存進細胞的「電池」ATP 裡。
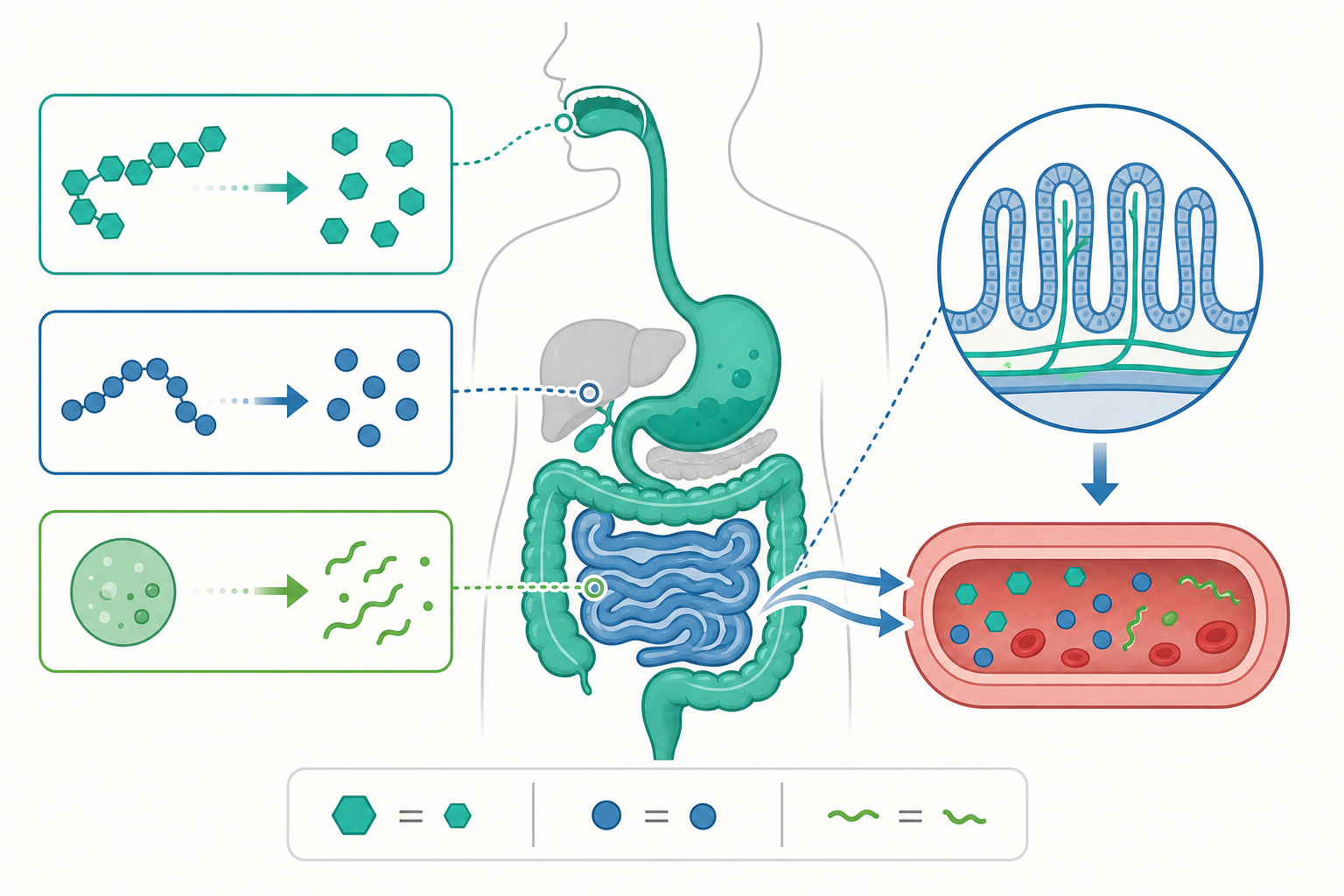
一趟食物的旅程:消化道導覽
讓我們跟著一口三明治,走一趟完整的消化道。這條從嘴巴到肛門的管子,全長約 9 公尺,比一輛公車還長,只是被巧妙地盤在你的肚子裡。
第一站:口腔。 牙齒把三明治磨碎(這是物理消化),唾液裡的澱粉酶開始把麵包裡的澱粉拆成較小的糖(這是化學消化)。你細嚼白飯久了會覺得有點甜,就是澱粉被拆成麥芽糖的緣故。
第二站:胃。 食物被吞下後,掉進像攪拌機一樣的胃。胃會分泌胃酸(鹽酸 HCl),把環境變得非常酸(pH 約 1.5–2),這個酸環境能殺菌,也能活化胃蛋白酶,開始拆解三明治裡火腿與起司的蛋白質。
第三站:小腸。 這是真正的主場。食物在這裡停留最久,分解得最徹底。胰臟送來各種消化酶(消化醣類、蛋白質、脂質的都有),肝臟分泌的膽汁則像洗碗精一樣,把大油滴「乳化」成小油滴,讓脂肪酶更容易作用。小腸內壁佈滿無數的「絨毛」,把吸收面積放大到約一個網球場那麼大——拆好的小分子,就從這裡被吸收進血液。
第四站:大腸。 剩下無法消化的纖維、水分和細菌,進入大腸。大腸主要負責回收水分,把剩餘的殘渣濃縮成糞便排出。
整條流程可以這樣對照:
| 器官 | 主要工作 | 處理對象 |
|---|---|---|
| 口腔 | 物理磨碎+澱粉酶 | 醣類 |
| 胃 | 胃酸+胃蛋白酶 | 蛋白質 |
| 小腸 | 多種酶+膽汁+吸收 | 醣、蛋白、脂質 |
| 大腸 | 回收水分 | 殘渣纖維 |
看不見的主角:酵素與表面積
如果沒有酵素(酶),消化會慢到無法維持生命。酵素是一種蛋白質催化劑,它能把化學反應加速數百萬倍,自己卻不會被消耗。每種酵素都很「挑食」,只認得特定的對象,這就是著名的「鑰匙與鎖」模型:澱粉酶這把鑰匙只開得了澱粉這把鎖,對蛋白質完全沒輒。
酵素還很在意「環境」。胃蛋白酶喜歡酸(在 pH 2 最有活力),但同一種酵素若進到中性的小腸就會失去作用;反過來,胰臟的酵素則偏好微鹼。這種精準的分工,讓不同營養素能在最適合的地點被處理。
另一個關鍵字是表面積。為什麼小腸要佈滿絨毛、絨毛上還有更小的微絨毛?因為吸收只發生在「表面」。一個簡單的對照:把一塊方糖丟進水裡很慢才溶,但把它磨成粉,瞬間就溶光了——同樣的體積,表面積越大,反應越快。我們的小腸用層層皺褶與絨毛,把吸收效率推到極致。
吃進去,不等於用得到
最後值得記住的一件事:「吃下去」和「被身體用到」是兩回事。 食物必須先被拆成夠小的分子,才能穿過小腸壁進入血液;進入血液後,還要被運送到各個細胞、進入細胞裡被代謝,才算真正派上用場。中間任何一個環節出問題——例如缺少某種消化酶(像乳糖不耐症缺乏乳糖酶),食物就會原封不動地通過,無法被利用,甚至引起不適。
所以「營養」不只是「吃什麼」,更是「身體能不能把它拆開、搬進去、用得上」。理解了消化與吸收,你就理解了食物變成你的全過程。
深入探討(研究所視角)
在分子層次,營養素的跨膜運輸遠比「吸收進血液」這句話複雜。以葡萄糖為例,小腸上皮細胞同時依賴兩套運輸蛋白:頂端膜的 SGLT1 利用鈉離子的電化學梯度進行次級主動運輸(鈉與葡萄糖同向共運,sodium-glucose cotransport),把葡萄糖逆濃度梯度泵入細胞;而基底側膜的 GLUT2 則以促進性擴散將葡萄糖順梯度送入血液。維持這套機制能量的根源,是基底膜上的 $\text{Na}^+/\text{K}^+$-ATPase,它水解 ATP 不斷把鈉打出細胞,重建鈉梯度——這正是口服補液鹽(ORS)治療脫水的生理基礎:在水中同時加入鈉與葡萄糖,能藉 SGLT1 共運帶動水分吸收。
酵素催化也可用嚴謹的動力學模型描述。米氏方程(Michaelis–Menten equation)刻畫了反應速率與受質濃度的關係:
$$v = \frac{V_{max}[S]}{K_m + [S]}$$
其中 $V_{max}$ 為最大反應速率,$K_m$(米氏常數)為達到半最大速率時的受質濃度,可反映酵素與受質的親和力——$K_m$ 越小,親和力越高。這個 1913 年提出的模型至今仍是酵素學、藥物代謝與臨床生化的基石。
近年的研究前沿,把「營養」從單一個體擴展到一整個生態系。腸道菌相(gut microbiota)藉由高通量定序與總體基因體學(metagenomics)被系統性解析後,人們發現腸道細菌能發酵宿主無法消化的膳食纖維,產生短鏈脂肪酸(如丁酸、丙酸、乙酸),這些代謝物不僅供應結腸細胞能量,還參與免疫調節與腸腦軸(gut-brain axis)的訊號傳遞。配合單細胞 RNA 定序(scRNA-seq),研究者已能解析腸道上皮中吸收細胞、杯狀細胞、腸內分泌細胞等不同類型的轉錄狀態與分化軌跡。
代謝層次的整合也走向系統生物學:以基因組尺度代謝模型(genome-scale metabolic model)搭配通量平衡分析(flux balance analysis, FBA),研究者能在電腦中模擬營養素在代謝網絡中的流向,預測在不同飲食或基因缺陷下細胞的代謝重分配。而 CRISPR-Cas9 基因編輯則讓人能精準剔除特定運輸蛋白或酵素基因,驗證其在養分吸收與代謝疾病(如糖尿病、肥胖)中的因果角色。從一口三明治到全身的代謝網絡,營養與消化的研究,正把生理學、分子生物學、微生物生態與計算模型編織成同一張地圖。