
從骨架到馬達:細胞骨架與細胞運動的分子機制
聚合動力學、馬達蛋白化學機械循環與細胞爬行的量化模型
從骨架到馬達:細胞為何能動
細胞看似柔軟,內部卻撐著一座持續拆解與重建的「動態鷹架」。這座鷹架不僅維持形狀,更把化學能轉換成機械力,讓細胞爬行、分裂、運送貨物。要理解細胞運動,不能停在「肌動蛋白、微管、中間絲」三大類的名詞層次,而必須進入聚合動力學、馬達蛋白的化學機械循環,以及力學回饋這三個量化層面。本文聚焦這些機制,並用幾個方程式把直覺釘成可計算的模型。
細胞骨架的核心矛盾在於:它既要夠穩定以承重,又要夠不穩定以快速重塑。這個矛盾的解,藏在核苷酸水解(ATP/GTP)為聚合提供的「方向性」與「非平衡」之中。
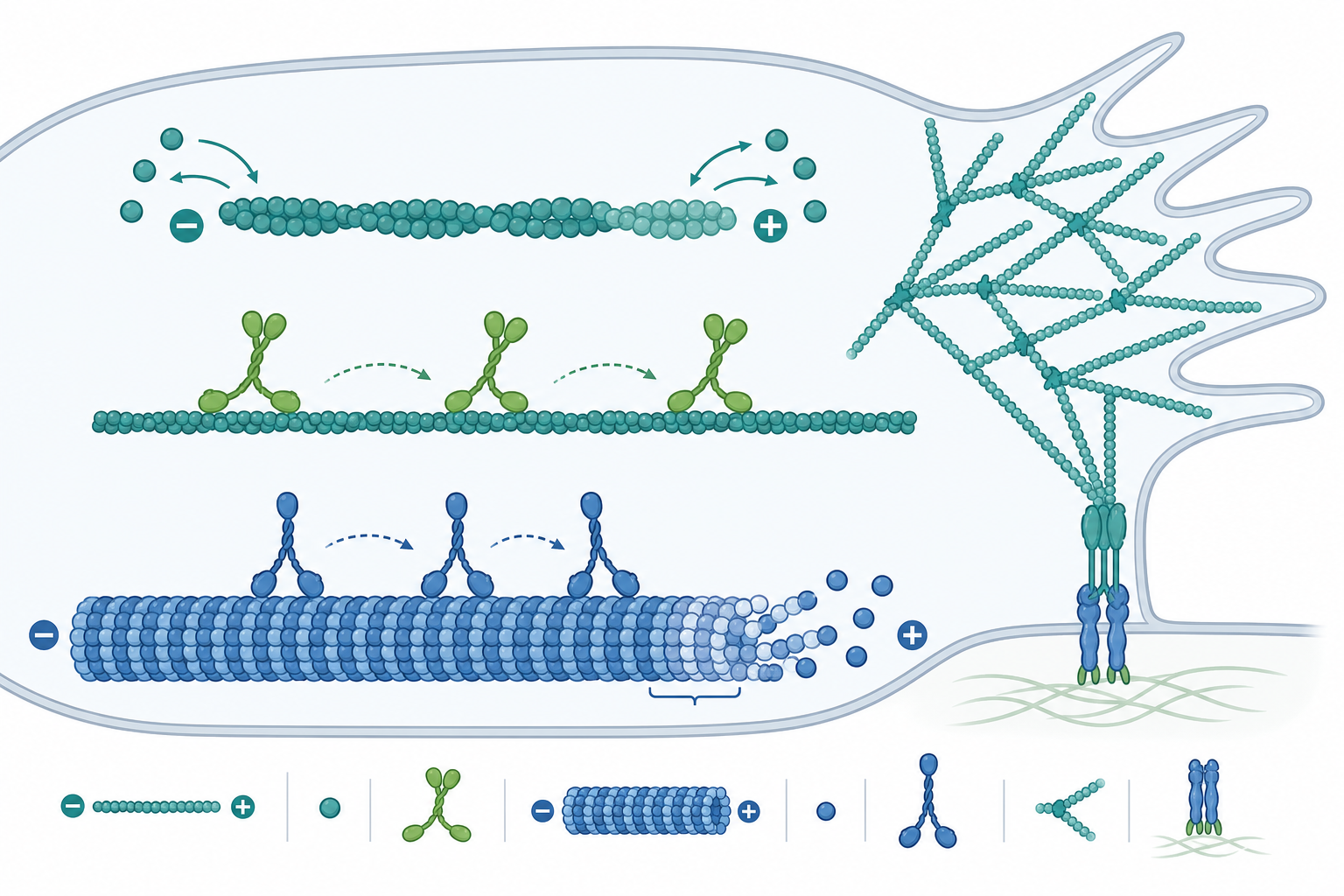
聚合動力學:踏車現象與動態不穩定
肌動蛋白絲(F-actin)與微管都是極性聚合物,兩端生長速率不同。以肌動蛋白為例,單體加入端的速率可寫成淨增長率:
$$\frac{dL}{dt} = k_{on}[\text{G-actin}] - k_{off}$$
其中 $k_{on}$ 為結合速率常數、$k_{off}$ 為解離速率常數,$[\text{G-actin}]$ 為游離單體濃度。當淨增長為零時,定義臨界濃度 $C_c = k_{off}/k_{on}$。關鍵在於:絲的兩端(plus 端、minus 端)各有不同的 $C_c$,因為 ATP-actin 與 ADP-actin 的親和力不同。當游離單體濃度落在兩端臨界濃度之間時,plus 端淨聚合、minus 端淨解聚,單體像輸送帶一樣由一端流向另一端——這就是踏車現象(treadmilling)。整條絲長度可維持恆定,但組成單體不斷被替換,這種「穩態卻持續耗能」正是非平衡系統的標誌。
微管則展現更極端的動態不穩定(dynamic instability):GTP-tubulin 在 plus 端形成「GTP 帽」,維持生長;一旦水解追上生長、帽消失,微管便瞬間崩解(catastrophe),縮回後又可能重新獲得帽而救援(rescue)。Mitchison 與 Kirschner 在 1984 年提出此模型,解釋了為何同一群微管在同一濃度下,有的在長、有的在縮。這種隨機切換讓微管能高效「搜尋」空間,例如在有絲分裂時捕捉著絲點。
馬達蛋白的化學機械循環
骨架提供軌道,真正產生位移的是馬達蛋白:沿肌動蛋白行走的肌凝蛋白(myosin)、沿微管行走的驅動蛋白(kinesin,朝 plus 端)與動力蛋白(dynein,朝 minus 端)。它們把 ATP 水解的自由能轉成構形改變,再轉成步進。
ATP 水解的自由能可寫成:
$$\Delta G = \Delta G^{\circ\prime} + RT \ln \frac{[\text{ADP}][\text{P}_i]}{[\text{ATP}]}$$
在細胞生理條件下(ATP 遠高於平衡值),$\Delta G$ 約為 $-50$ 至 $-60\ \text{kJ/mol}$。Kinesin-1 每水解一個 ATP 前進約 $8\ \text{nm}$(恰為微管 αβ-tubulin 異二聚體的週期),因此其機械輸出上限約:
$$F_{stall} = \frac{|\Delta G|}{d} \approx \frac{55{,}000\ \text{J/mol}}{6.022\times10^{23}\ \text{mol}^{-1} \times 8\times10^{-9}\ \text{m}} \approx 11\ \text{pN}$$
實測 kinesin 的停滯力(stall force)約 $5\text{–}7\ \text{pN}$,低於此理論上限,差距正是因為循環並非 100% 效率,且部分能量耗散於熱與步進的隨機性。
馬達的行走速率對 ATP 濃度呈飽和關係,可用 Michaelis–Menten 形式描述步進率:
$$v = \frac{v_{max}[\text{ATP}]}{K_M + [\text{ATP}]}$$
定量小範例:假設某 kinesin 的 $v_{max} = 800\ \text{nm/s}$、$K_M = 50\ \mu\text{M}$,細胞內 $[\text{ATP}] = 2\ \text{mM} = 2000\ \mu\text{M}$。代入:
$$v = \frac{800 \times 2000}{50 + 2000} = \frac{1{,}600{,}000}{2050} \approx 780\ \text{nm/s}$$
由於 $[\text{ATP}] \gg K_M$,馬達幾乎以最高速行走(達 $v_{max}$ 的 97.6%)。若 ATP 降到 $50\ \mu\text{M}$(即 $K_M$),速率恰為 $v_{max}$ 的一半($400\ \text{nm/s}$),這正是 $K_M$ 的物理意義。
細胞爬行:力的產生與回饋
爬行細胞(如纖維母細胞、免疫細胞)的運動可拆成四步:前緣突出(protrusion)、黏附(adhesion)、收縮(contraction)、後緣脫附(de-adhesion)。前緣的片狀偽足(lamellipodium)由 Arp2/3 複合體催化的分枝肌動蛋白網絡推動。Brownian ratchet(布朗棘輪)模型指出:膜的熱漲落短暫讓出空間,肌動蛋白單體趁隙插入絲端,把膜「卡」在新位置,於是熱運動被整流成定向推力。
收縮力來自非肌肉肌凝蛋白 II 在應力纖維上的滑動,並透過黏著斑(focal adhesion)把力傳到細胞外基質。這裡有個重要的力學回饋:黏著斑是機械敏感(mechanosensitive)的,受力時會招募更多蛋白而增強,這由 talin 在張力下展開、暴露 vinculin 結合位點所介導。基質硬度因此能調控細胞行為——細胞會朝較硬的基質遷移(durotaxis)。整體遷移可視為前緣聚合速率、馬達收縮力與黏附壽命三者競爭下的淨結果。
深入探討(研究所視角)
前述模型多半是「平均場」描述,但近十年的結構生物學、單分子與系統生物學把細胞運動推進到更嚴謹的層次。
結構生物學:冷凍電鏡與構象捕捉。 低溫電子顯微術(cryo-EM)的解析度革命,讓研究者在近原子尺度看見馬達在軌道上的各個構象狀態。微管 plus 端「GTP 帽」的結構特徵——GTP-tubulin 處於相對伸直的晶格、水解後晶格收縮並產生彎曲應變——支持了「儲存的機械應變驅動崩解」的觀點。Cryo-electron tomography(冷凍電子斷層)更能在接近原生的細胞切片中重建肌動蛋白網絡的三維分枝幾何,量測分枝角約 70 度,直接驗證 Arp2/3 的幾何模型。dynein 的力臂(linker)在不同核苷酸態下的擺動,也是靠結構快照才得以連成連續的動力衝程(power stroke)。
單分子與光鉗。 光鑷(optical tweezers)能在皮牛頓尺度測量單一馬達的步進、停滯力與外力下的滯留時間,揭示其化學機械耦合對負載的依賴:外力增大時,kinesin 的離軌率上升、步進變慢,符合受力依賴的反應速率(類 Bell 模型 $k(F) = k_0 \exp(F x^\ddagger / k_B T)$,其中 $x^\ddagger$ 為過渡態的特徵距離)。這類實驗顯示馬達並非剛性齒輪,而是受熱噪聲與外力共同調制的隨機機器。
主動物質與系統層級模型。 細胞骨架是「主動物質(active matter)」的範式:馬達持續注入能量,使系統處於非平衡穩態,能自發產生流動、漩渦與拓撲缺陷。以肌動蛋白—肌凝蛋白為基礎的重構系統(reconstituted cortex)顯示,調節馬達密度與交聯比例即可在「均勻收縮」與「圖案化流動」間切換。理論上,這些可用主動凝膠(active gel)連續介質方程描述,把應力張量拆成被動黏彈項與正比於序參數的主動項。
單細胞與多體學整合。 單細胞 RNA-seq 與蛋白體學讓研究者把遷移表型對應到調控網絡:Rho GTPase 家族(RhoA、Rac1、Cdc42)的時空活化模式決定突出、黏附與收縮的協調。活細胞 FRET 生物感測器揭示 Rac1 與 RhoA 在前緣與後緣形成互斥的活性梯度,構成自組織的極性回饋迴路。把這些訊號動力學嵌入反應—擴散模型,能再現細胞自發極化與趨化轉向。
近期方向。 條件性蛋白降解(如 auxin-inducible degron)與光遺傳學讓研究者得以在秒級時間尺度急性擾動特定骨架成分,分辨「直接效應」與「代償適應」;而把 cryo-ET 的結構參數餵入分子動力學與粗粒化模擬,正逐步把「單一分子的力學」與「整個細胞皮層的湧現行為」連成一條可計算的多尺度鏈。細胞運動因此不再只是描述性現象,而是一個能從自由能、步進距離、聚合速率到群體流動逐層推導的量化問題。