
從行為到基因:行為生態學與保育遺傳的量化整合
最適化、性擇、有效族群大小與遺傳救援——保育為何必須同時讀懂行為策略與等位基因頻率
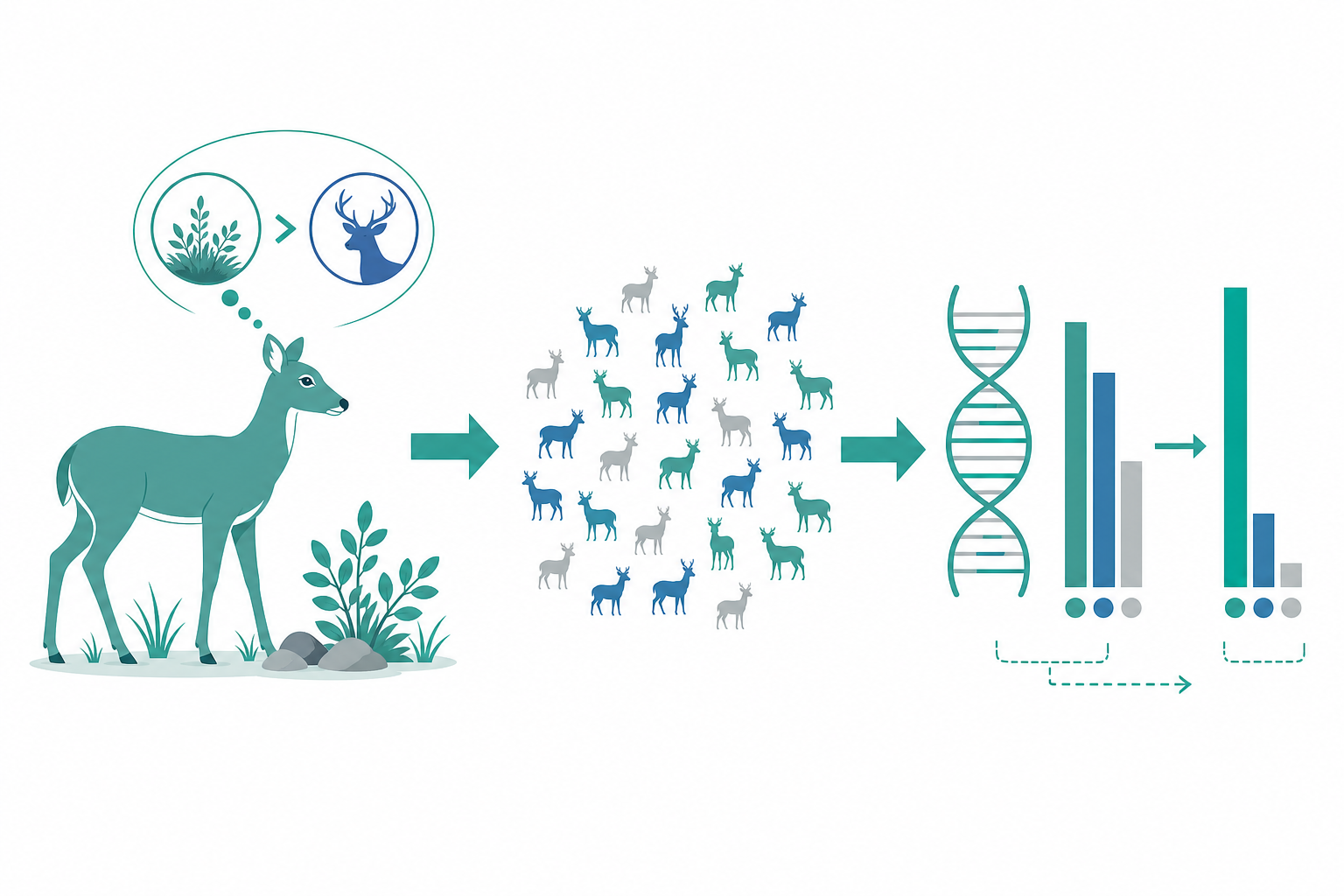
從個體行為到族群命運:為什麼保育需要懂行為與基因
一頭公象在乾旱季記得三十年前的水源、一隻雌鳥對親緣訊號的辨識、一群魚在掠食壓力下的聚散——這些看似瑣碎的行為決策,最終都會寫進族群的等位基因頻率裡。行為生態學研究行為如何在天擇下被塑造成適應策略;保育遺傳則追問當族群縮小、隔離、近交時,這些策略還能不能維繫遺傳變異與演化潛力。兩者交會處,正是現代保育生物學最吃力也最關鍵的戰場:我們保護的不只是個體數量,而是一套能持續演化的「行為—基因」共構系統。
本篇假設你已掌握天擇、孟德爾遺傳與基本族群生態,直接進入機制與量化層次。
行為作為適應:最適化與賽局的量化骨架
行為生態學的核心假設是行為表現型受天擇塑造,趨向使適應度最大化的策略。最經典的量化工具是 最適覓食理論(Optimal Foraging Theory)。以邊際值定理(Marginal Value Theorem)為例,覓食者應在某資源斑塊的瞬時收益率下降到「環境平均收益率」時離開。設斑塊累積能量為 $g(t)$、斑塊間移動時間為 $\tau$,則最適停留時間 $t^*$ 滿足
$$g'(t^*) = \frac{g(t^*)}{\tau + t^*}$$
也就是離開瞬間的邊際收益率等於整體平均收益率。這條切線條件可由作圖直接求解,是大量野外取食數據的理論錨點。
當個體的最適策略取決於他人怎麼做時,最適化要升級為 演化穩定策略(ESS, Maynard Smith)。在 Hawk–Dove 賽局中,若爭奪資源價值為 $V$、受傷成本為 $C$ 且 $C>V$,則混合 ESS 的鷹策略比例為 $p^* = V/C$。這解釋了為何純粹好鬥不會席捲族群——攻擊性是頻率依賴下的平衡,而非單純「越兇越好」。這類頻率依賴選擇也是維持行為多型(如顯性與衛星交配策略)的機制之一。
性擇、訊號與「好基因」假說
性擇是行為最劇烈塑造表現型的力量。Zahavi 的 累贅原則(Handicap Principle) 主張誇耀性訊號之所以誠實,正因其成本高昂:只有高品質個體負擔得起昂貴的尾羽或鳴唱,使訊號難以作弊。Grafen 後續以賽局模型證明,當訊號成本與個體品質呈負相關(高品質者邊際成本較低)時,誠實訊號可成為演化穩定的訊號均衡。
這對保育的意涵在於:性擇往往依賴 遺傳變異。若「好基因」假說成立,雌性偏好的訊號反映了抗病、代謝等位點的優劣,而這正是被近交與遺傳漂變最先侵蝕的部分。MHC(主要組織相容性複合體)的擇偶偏好就是典型案例——許多物種偏好 MHC 相異的配偶,以維持後代免疫多樣性。族群一旦縮小,MHC 多樣性流失,連帶削弱基於嗅覺或免疫訊號的擇偶行為。
保育遺傳的量化基準:Hardy–Weinberg 與有效族群大小
保育遺傳的所有偏離,都以 Hardy–Weinberg 平衡 為對照基準。對雙等位基因座,基因型頻率為
$$p^2 + 2pq + q^2 = 1, \quad p + q = 1$$
真實小族群會系統性偏離此平衡,原因正是近交與漂變。近交係數 $F$ 衡量觀測異型合子相對期望的虧缺:
$$F = \frac{H_e - H_o}{H_e}$$
其中 $H_e = 2pq$ 為期望異型合子率、$H_o$ 為觀測值。$F>0$ 代表近交累積。
更核心的是 有效族群大小 $N_e$。遺傳漂變導致每世代雜合度的流失率約為
$$H_{t} = H_0\left(1 - \frac{1}{2N_e}\right)^{t}$$
關鍵在於 $N_e$ 幾乎總是遠小於普查族群數 $N_c$——而行為正是壓低 $N_e$ 的主因。一夫多妻、偏斜的繁殖成功、性別比失衡都會使 $N_e \ll N_c$。在性別比偏斜下:
$$N_e = \frac{4 N_m N_f}{N_m + N_f}$$
定量小範例: 一座保護區有 90 隻成體,但因強烈一妻多夫的交配系統,僅 5 隻雄性與 85 隻雌性實際參與繁殖。代入:
$$N_e = \frac{4 \times 5 \times 85}{5 + 85} = \frac{1700}{90} \approx 18.9$$
雖然普查數 90,有效族群僅約 19。代入漂變式,每世代雜合度流失約 $1/(2 \times 18.9) \approx 2.6\%$。十個世代後保留比例為 $(1-0.026)^{10} \approx 0.77$,即喪失近四分之一的中性遺傳變異。這正說明:忽略交配行為來估算遺傳健康,會嚴重高估族群的演化壽命。
近交衰退、適應潛力與族群生存力
近交衰退(inbreeding depression)使隱性有害等位基因暴露於同型合子,壓低存活與繁殖。著名的瑞典 Adder(北蝰)孤立族群與佛州山獅(Florida panther)案例都顯示,當 $N_e$ 跌破門檻,畸形精子、幼體死亡率上升等表現型隨之出現。保育上常引用的 50/500 法則——短期避免近交衰退需 $N_e \geq 50$,長期維持演化潛力需 $N_e \geq 500$——雖是粗略經驗值,但其量化精神正源自上述漂變與近交方程。
行為層面的對策包括 遺傳救援(genetic rescue):引入少數外來個體以注入新等位基因。佛州山獅引入德州山獅後,後代健康與族群數顯著回升,是行為(遷入與交配)直接逆轉遺傳衰退的範例。但成效取決於外來者能否被接納繁殖——若社會結構排斥外來個體,遺傳救援在分子層次再合理,行為層次仍可能失敗。
深入探討(研究所視角)
進入研究前沿,行為與保育遺傳的整合正被三股技術力量重塑:基因體尺度的選擇偵測、單細胞與多體學、以及機制性的數學模型。
保育基因體學與選擇掃描。 從少數微衛星座位邁向全基因體重定序後,研究者可直接量化 連鎖不平衡 推估的歷史 $N_e$,並以 runs of homozygosity(ROH) 計算個體基因體近交係數 $F_{ROH}$——長 ROH 段代表近期近交,短段反映遠古瓶頸。同時,跨族群的 $F_{ST}$ 與 整合單套型同源性(iHS)、XP-EHH 等統計可定位近期受正向選擇的位點。對行為而言,這意味著我們能反向追問:被天擇保留的,是否正是與覓食、社會性、抗病行為相關的基因座?例如 vasopressin/oxytocin 受體基因(如 avpr1a) 與田鼠(Microtus)配偶連結行為的關聯,已成為「行為基因座如何在族群結構中分佈」的範式系統。
單細胞與神經分子機制。 行為的近因(proximate cause)正被單細胞 RNA-seq 與空間轉錄體解析到神經元型別層次。社會行為(如優勢階層、親代照護)對應的 轉錄體可塑性 顯示,同一基因組可依社會情境快速切換表現型——這是「表現型可塑性如何緩衝遺傳變異流失」的分子證據。表觀遺傳修飾(DNA 甲基化、組蛋白修飾)則提供跨世代傳遞行為傾向的非序列途徑,挑戰了「遺傳變異是適應潛力唯一儲庫」的傳統假設。
系統生物學與整合族群模型。 最前沿的保育預測已從單一 $N_e$ 走向 整合族群模型(Integrated Population Models, IPM) 與 個體本位模型(IBM/ABM):將標放重捕、遺傳取樣、行為追蹤(GPS、加速度計生物紀錄)資料於貝氏框架下聯合估計。這類模型能捕捉 族群統計—遺傳回饋迴路——例如近交衰退降低存活率,存活率下降又進一步壓低 $N_e$,形成滅絕漩渦(extinction vortex)的正回饋。將 logistic 成長 $\frac{dN}{dt}=rN(1-N/K)$ 與隨機性、Allee 效應(低密度下適應度反降,常源於難以找到配偶的行為機制)耦合後,模型可輸出 最小存活族群(MVP) 的機率分佈而非單一閾值。
結構與分子演化的交會。 在分子層次,MHC 與免疫受體的 超基因(supergene) 與平衡選擇研究,正用結構生物學解釋為何某些等位基因被長期維持——抗原結合溝槽的胺基酸多型直接決定病原辨識廣度。將蛋白結構約束納入族群遺傳模型,使我們能預測哪些變異流失最致命。這條從 行為(擇偶、社會性)→ 神經分子機制 → 基因體選擇訊號 → 蛋白結構功能 的完整因果鏈,正是下一代保育遺傳試圖閉合的迴路:不再只問「還剩多少變異」,而是問「剩下的變異,能不能支撐這個物種繼續演化下去」。