
演化:一個簡單想法,如何解釋所有生命
天擇、變異與族群遺傳,串起從胡椒蛾到加拉巴哥雀鳥的生命統一故事。
演化:一個解釋所有生命的簡單想法
如果你抬頭看一片森林、低頭看腳邊的螞蟻,再想想浴室裡那些怎麼擦也擦不掉的黴菌,會覺得這些生命彼此差異實在太大。但生物學家有一個驚人的主張:所有這些生命,其實都來自同一個古老的祖先,靠著一個非常簡單的機制,慢慢「長」成今天的模樣。這個機制,就是演化。
達爾文(Charles Darwin)在《物種起源》裡提出的核心想法,可以用三句話講完:
- 同一群生物的個體之間,會有變異(有的跑得快、有的耐渴、有的顏色深)。
- 這些變異有一部分可以遺傳給後代。
- 環境資源有限,比較適應環境的個體,平均能留下比較多後代。
把這三件事放在一起,結果就是:能幫助生存與繁殖的特徵,會在世代之間越來越普遍。這個過程叫做天擇(natural selection)。請特別注意:天擇不是「最強者生存」,而是「最適合當下環境者,留下最多後代」。環境變了,誰算「適合」也會跟著變。
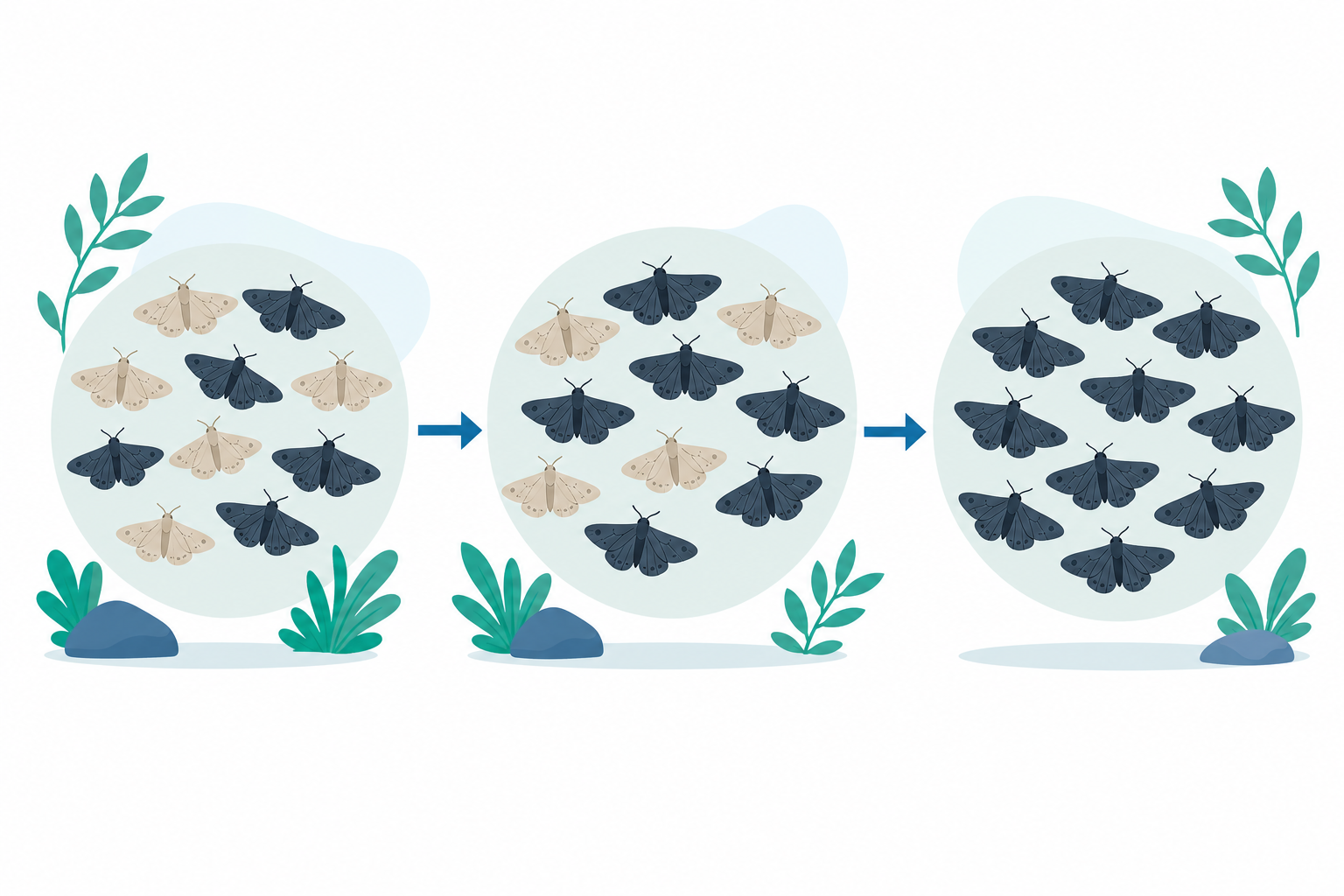
一個看得見的例子:胡椒蛾
英國工業革命時期有一種蛾叫胡椒蛾(peppered moth)。牠原本大多是淺色的,停在長滿淺色地衣的樹皮上,鳥幾乎看不到牠。但工廠燒煤把樹皮燻黑、地衣也死了,淺色蛾反而變得超級顯眼,一隻接一隻被鳥吃掉。
這時候原本罕見的深色變異蛾就佔了便宜:牠們停在黑樹皮上不容易被發現,活得久、生得多。短短幾十年,工業區的胡椒蛾族群就從「幾乎全淺色」變成「大多深色」。後來空氣污染改善、樹皮恢復,淺色蛾又慢慢回來了。
這個例子漂亮在哪裡?它讓我們看見:演化不是蛾「想要」變黑,也不是單一隻蛾在一生中變色。變異原本就存在(深色基因一直都在族群裡),天擇只是改變了各種變異在族群中的比例。換句話說——
演化的主角不是個體,而是族群。個體會生老病死,但「族群中某個基因有多常見」這件事,才是演化真正在改變的東西。
演化的原料:變異從哪裡來?
天擇只能從現有的變異裡「挑選」,它沒辦法無中生有。那變異本身從哪來?主要有兩個來源:
- 突變(mutation):DNA 在複製時偶爾出錯,產生新的版本(等位基因)。突變是隨機的,多數沒影響、少數有害、極少數剛好有利。但正是這些罕見的有利突變,提供了演化最原始的材料。
- 有性生殖的重組:父母的基因在製造精卵時重新洗牌,讓後代有全新的組合。這也是為什麼兄弟姊妹長得不一樣。
除了天擇,還有兩個力量會改變族群的基因比例:基因流(gene flow),也就是個體在族群間遷移、把基因帶來帶去;以及遺傳漂變(genetic drift),在小族群裡,純粹因為「運氣」,某些基因可能憑機率就消失或變多——就像丟硬幣,丟 4 次很容易全是正面,但丟 4000 次幾乎不可能。族群越小,運氣的影響越大。
怎麼知道族群「正在」演化?哈溫平衡
生物學家需要一把尺,來判斷一個族群到底有沒有在演化。這把尺叫哈溫平衡(Hardy–Weinberg equilibrium)。它描述一個「理想中完全沒有演化」的族群,基因比例會怎麼分布。
假設某基因只有兩個版本 $A$ 和 $a$,設 $A$ 的頻率為 $p$、$a$ 的頻率為 $q$,因為只有這兩種,所以:
$$p + q = 1$$
那麼下一代各種基因型的比例就是:
$$p^2 + 2pq + q^2 = 1$$
其中 $p^2$ 是 $AA$、$2pq$ 是 $Aa$、$q^2$ 是 $aa$。
舉個具體例子:假設 $A$ 的頻率 $p = 0.7$,$a$ 的頻率 $q = 0.3$。代入計算:
- $AA$ 的比例 $= p^2 = 0.7^2 = 0.49$(49%)
- $Aa$ 的比例 $= 2pq = 2 \times 0.7 \times 0.3 = 0.42$(42%)
- $aa$ 的比例 $= q^2 = 0.3^2 = 0.09$(9%)
加起來 $0.49 + 0.42 + 0.09 = 1$,剛好 100%。
關鍵在於:如果一個族群真的沒有演化(沒有突變、沒有天擇、沒有遷移、沒有漂變、隨機交配),那一代又一代,這些比例都不會變。所以反過來——只要我們實際觀察到比例偏離了哈溫平衡的預測,就代表上述某個演化力量正在作用。哈溫平衡因此不是描述真實世界,而是當作一個「對照基準線」,讓偏差現身。
物種是怎麼分出來的?
如果一個族群被分成兩半(比方一條河、一座新長出來的山把牠們隔開),兩邊各自累積不同的突變、面對不同的環境、被不同方向的天擇拉扯,久而久之差異越來越大。直到有一天,就算把牠們放回一起,也已經無法成功交配繁殖出有生育能力的後代——這時我們就說,一個物種變成了兩個物種,這個過程叫物種形成(speciation)。
加拉巴哥群島上達爾文觀察到的那些雀鳥,喙的形狀各不相同,正是這樣分化出來的:有的喙厚短適合咬硬種子、有的尖細適合捉蟲。同一個祖先,因為各島環境不同,被天擇雕成了不同的樣子。
深入探討(研究所視角)
入門段把天擇講成「比例的改變」,但現代演化生物學的核心,其實是把這套邏輯數學化、分子化的「演化綜合論(Modern Synthesis)」與其後的擴展。
在族群遺傳學的形式架構裡,天擇的強度以選擇係數(selection coefficient) $s$ 量化,個體的適應度(fitness) $w = 1 - s$。對於一個頻率為 $q$ 的等位基因,在加性選擇下其單代變化可近似為:
$$\Delta q \approx \frac{s \, pq}{\bar{w}}$$
其中 $\bar{w}$ 為族群平均適應度。這個式子揭示一個非直覺的結論:當有害等位基因已經很罕見($q$ 很小)時,$\Delta q$ 也趨近於零——這正是為什麼天擇難以徹底清除隱性有害基因,它們得以低頻率長期潛伏。木村資生(Motoo Kimura)的中性理論(Neutral Theory)進一步主張,分子層次上多數的變異其實是「選擇中性」的,命運由遺傳漂變而非天擇主導;漂變的強度與有效族群大小(effective population size) $N_e$ 成反比,這把 $N_e$ 推上了演化推論的核心位置。
分子機制上,今天我們不只看「基因頻率」,更看基因組架構如何約束演化路徑:基因重複(gene duplication)提供冗餘副本,讓其中一份得以在不喪失原功能的情況下累積突變、演化出新功能(neofunctionalization);順式調控元件(cis-regulatory element)的改變往往比蛋白質編碼序列的改變更能驅動形態演化,因為它能在特定時空調整基因表現而不牽動全身——這是「演化發育生物學(evo-devo)」的中心洞見。
技術前沿正在改寫我們能問的問題。單細胞與單分子定序讓研究者能在同一個體內追蹤體細胞突變的譜系,重建腫瘤或胚胎發育中的「微演化」;古 DNA(ancient DNA) 定序則讓我們直接讀取尼安德塔人等已滅絕族群的基因組,量化基因流而非僅憑骨骼推測。CRISPR-Cas9 不只是編輯工具,它本身就是細菌對抗噬菌體的演化軍備競賽產物——一套可遺傳的適應性免疫系統;而以 CRISPR 為基礎的基因驅動(gene drive),更讓人類有能力刻意偏移野生族群的孟德爾遺傳比例,引發深刻的生態與倫理辯論。
最後,演化與其他維度的連結正日益緊密。在系統生物學視角下,適應度地形(fitness landscape)上的路徑受到基因間上位效應(epistasis) 的強烈約束,並非每條向上的路都走得通;而實驗演化——如 Lenski 的大腸桿菌長期演化實驗,已連續培養超過七萬代——讓「演化」從歷史推論變成可在實驗室即時觀測、即時定序的活科學。