
消化吸收的神經與激素調控:從質子幫浦到腦腸軸的量化機制
三相分泌、Hill 協同飽足訊號、米氏轉運動力學,與單細胞時代的腸腦突觸
從一口食物說起:腸道是一座會自己決策的化學工廠
當你吞下一口食物,腸道並不是被動地等著消化酶來分解它。事實上,在食物還沒抵達之前,胃酸就已經開始分泌;在養分吸收的同時,飽足訊號正悄悄上傳腦幹。腸道擁有約 $2$ 至 $6$ 億個神經元(與一隻貓的整個中樞神經系統相當),加上數十種胜肽激素,構成一套能在毫秒到小時尺度上自主調控分泌、蠕動與吸收的「第二大腦」。本篇要談的,正是這套神經與激素調控網路如何在分子層次協調消化吸收,並嘗試用幾個量化模型刻畫它的行為。
直覺上我們把消化想成「機械研磨+酶解」,但真正決定效率的,是訊號的時序與劑量。同樣的一餐,若飽足訊號提早或延遲幾分鐘釋放,攝食量可能相差數百大卡。理解這套調控,才是理解營養吸收的關鍵。
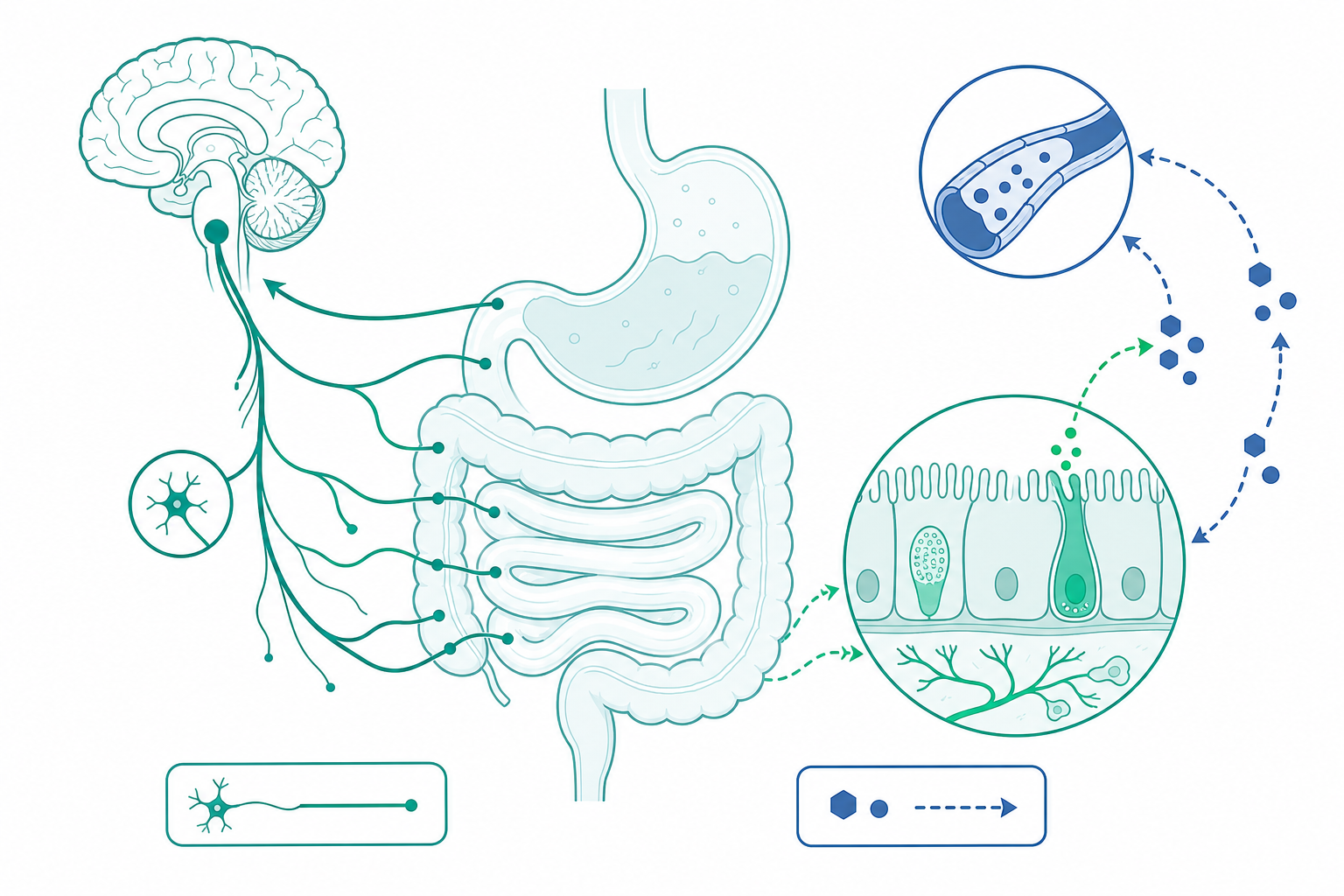
腦腸軸與三相分泌:頭期、胃期、腸期
消化的調控傳統上分為三相,三者在時間上重疊而非接續。
頭期(cephalic phase)由視覺、嗅覺、咀嚼經迷走神經(vagus nerve)的副交感纖維觸發,迷走節後神經元釋放乙醯膽鹼,刺激胃壁細胞(parietal cell)分泌鹽酸、主細胞分泌胃蛋白酶原。這一相佔總胃酸分泌約 $30\%$,且在食物入胃前即已啟動,是典型的前饋(feed-forward)控制。
胃期(gastric phase)由胃的機械牽張與蛋白質消化產物(胺基酸、胜肽)觸發。G 細胞釋放胃泌素(gastrin),經血流回到壁細胞放大酸分泌,同時局部組織胺由 ECL 細胞釋放,透過 $\mathrm{H_2}$ 受體進一步增益。三條路徑(神經、內分泌、旁分泌)在壁細胞的質子幫浦 $\mathrm{H^+/K^+}$-ATPase 上匯流,形成乘性放大。
腸期(intestinal phase)則以負回饋為主軸。當酸性食糜進入十二指腸,S 細胞分泌胰泌素(secretin)刺激胰臟與膽管分泌碳酸氫鹽中和酸;I 細胞偵測脂肪酸與胺基酸後分泌膽囊收縮素(CCK),促進胰酶分泌與膽囊收縮,並延緩胃排空。這套「腸抑胃」(enterogastrone)機制確保上游不會傾倒超過下游處理能力的食糜。
質子幫浦與壁細胞:一個熱力學受限的離子泵
壁細胞分泌鹽酸是全身最極端的離子梯度建立過程之一。胃腔內 $\mathrm{pH}$ 可低至 $0.8$,相對於細胞內 $\mathrm{pH} \approx 7.2$,跨膜質子濃度差達 $10^{6}$ 倍以上。
我們可以用能斯特方程式(Nernst equation)估算維持此梯度所需的電化學功。對 $\mathrm{H^+}$ 而言,平衡電位為
$$E_{\mathrm{H^+}} = \frac{RT}{zF} \ln \frac{[\mathrm{H^+}]_{\text{out}}}{[\mathrm{H^+}]_{\text{in}}}$$
取 $T = 310\,\mathrm{K}$、$z = +1$,$\dfrac{RT}{F} \approx 26.7\,\mathrm{mV}$。若跨膜質子濃度比為 $10^{6}$,則
$$E_{\mathrm{H^+}} = 26.7 \times \ln(10^{6}) = 26.7 \times 13.8 \approx 369\,\mathrm{mV}$$
換算為每莫耳質子所需自由能 $\Delta G = zF E \approx 96\,485 \times 0.369 \approx 35.6\,\mathrm{kJ/mol}$。這正解釋了為何 $\mathrm{H^+/K^+}$-ATPase 需要 ATP 水解($\Delta G \approx -50\,\mathrm{kJ/mol}$)來驅動——一個 ATP 約可推送一至二個質子逆梯度,逼近熱力學上限。臨床上的氫離子幫浦抑制劑(PPI,如 omeprazole)正是不可逆共價封閉此酶,效果持久。
飽足激素的劑量—反應:用 Hill 方程描述
進食後的飽足感由多種激素共同編碼,其中 CCK 與 GLP-1(類升糖素胜肽-1)作用於迷走傳入神經與後腦。受體結合往往呈現協同性(cooperativity),可用 Hill 方程描述受體佔據率 $\theta$:
$$\theta = \frac{[L]^{n}}{K_d^{\,n} + [L]^{n}}$$
其中 $[L]$ 為激素濃度、$K_d$ 為解離常數、$n$ 為 Hill 係數。當 $n > 1$ 時曲線呈 S 形(sigmoidal),意味著存在「開關式」的閾值效應——飽足訊號在某濃度附近急遽放大,這在生理上有助於明確終止進食,而非緩慢拖延。
定量小範例。 假設某 CCK 受體 $K_d = 5\,\mathrm{nM}$、$n = 2$,問餐後 $[L] = 10\,\mathrm{nM}$ 時受體佔據率為何?
代入:
$$\theta = \frac{10^{2}}{5^{2} + 10^{2}} = \frac{100}{25 + 100} = \frac{100}{125} = 0.80$$
即 $80\%$ 受體被佔據。若改用 $n = 1$(無協同):$\theta = 10/(5+10) = 0.667$,僅 $67\%$。可見協同性 $n=2$ 讓相同濃度產生更陡的飽足反應。若把濃度從 $5\,\mathrm{nM}$ 升到 $10\,\mathrm{nM}$,$n=2$ 的佔據率由 $0.50$ 跳到 $0.80$(增加 $0.30$),而 $n=1$ 僅由 $0.50$ 升到 $0.667$(增加 $0.17$)——這就是協同性帶來的「開關銳度」。
養分吸收的酶動力學與轉運上限
吸收不只是擴散。小腸刷狀緣的雙醣酶水解與膜轉運蛋白都遵循飽和動力學,可用米氏方程式(Michaelis–Menten)刻畫反應速率:
$$v = \frac{V_{\max}[S]}{K_m + [S]}$$
以 $\mathrm{SGLT1}$(鈉—葡萄糖共轉運蛋白)為例,其對葡萄糖的 $K_m$ 約在 $0.5$–$2\,\mathrm{mM}$ 之間。當腸腔葡萄糖濃度遠高於 $K_m$ 時,轉運趨近 $V_{\max}$ 而飽和;此時神經與激素調控(如 GLP-1 延緩胃排空、降低餐後血糖峰值)反而成為決定吸收速率的上游瓶頸。換言之,激素藉由控制「上料速度」來避免轉運系統超載,這正是腸促胰素(incretin)軸的生理意義:GLP-1 與 GIP 在養分入腸時提前刺激胰島素分泌,使血糖更平滑。
蠕動的節律:Cajal 間質細胞與慢波
消化道平滑肌並非隨機收縮,而是由 Cajal 間質細胞(ICC)作為節律起搏器,產生每分鐘約 $3$ 次(胃)到 $12$ 次(十二指腸)的慢波(slow wave)。腸神經系統(ENS)的肌間神經叢協調收縮節段間的相位差,形成由口向肛的推進蠕動。迷走與交感輸入則像「音量旋鈕」,調節而非主導這套內生節律——即使切斷外來神經,腸道仍能維持基本蠕動,這是 ENS 自主性的直接證據。
深入探討(研究所視角)
主體談的是經典三相與激素—受體層次的調控。最前沿的問題,則指向訊號的細胞解析度與系統整合。
腸內分泌細胞的「感官」分子身分。 過去以為腸內分泌細胞(enteroendocrine cell, EEC)只是被動的化學偵測器,但近十年單細胞 RNA 定序顯示,同一型 EEC 可同時表現多種激素(CCK、GLP-1、PYY 共表現),傳統「一細胞一激素」教條被推翻,取而代之的是依空間位置與管腔內容物動態切換分泌譜的「彈性身分」模型。更關鍵的是 2018 年起確立的神經足細胞(neuropod cell)——某些 EEC 具有突觸樣的基底延伸,與迷走傳入神經形成單一突觸連結,可用麩胺酸作為快速神經傳遞物,將管腔糖訊號在毫秒級而非分鐘級傳遞至腦幹。這把「腸—腦」從緩慢的內分泌通訊升格為真正的突觸電路。
營養感測受體的結構生物學。 甜味受體 T1R2/T1R3、脂肪酸受體 GPR40/GPR120、胺基酸感測的 CaSR 與 GPRC6A 皆屬 class C GPCR。冷凍電顯(cryo-EM)近年解出多個 class C GPCR 全長結構,揭示其特徵的 Venus flytrap 配體結合域與七次跨膜域之間的變構耦合機制。理解這些結構,是設計選擇性 GLP-1 分泌促進劑的基礎——這也與當前 GLP-1 受體促效劑(如 semaglutide)的臨床熱潮直接相關,後者透過延緩胃排空與中樞飽足訊號顯著降低食慾。
系統生物學與計算模型。 餐後代謝的整體行為已可用常微分方程系統建模,例如把胃排空、葡萄糖吸收、incretin 分泌、胰島素—升糖素回饋耦合成多區室模型,用以擬合口服葡萄糖耐量試驗(OGTT)曲線。這類模型常引入希爾型飽和項與時間延遲(time-delay)項來捕捉胃排空的非線性。延伸到族群層次,飽足訊號與攝食的負回饋甚至可借用 logistic 形式 $\dfrac{dN}{dt} = rN\left(1 - \dfrac{N}{K}\right)$ 的數學直覺——攝食量趨近某「承載量」後自我抑制,雖然生理載體不同,但收斂式回饋的數學骨架相通。
菌—腸—腦軸的調控新層。 腸道微生物代謝產物(短鏈脂肪酸如丁酸、丙酸)可活化 EEC 上的 FFAR2/FFAR3 受體,調節 GLP-1 與 PYY 分泌;膽汁酸經 TGR5 受體亦參與此回路。這意味著激素調控網路其實是三方對話:宿主神經、宿主內分泌、與微生物代謝物共同設定分泌的基線與增益。如何在系統生物學模型中正確納入這層擾動,仍是開放問題,也是未來精準營養(precision nutrition)的核心戰場。