
光呼吸與 C4/CAM 的演化適應:Rubisco 缺陷的補救工程
從專一性因子、碳濃縮泵到結構約束與作物工程的量化視角
光呼吸:Rubisco 的「先天缺陷」與演化的補救工程
植物把光當能量、把二氧化碳變成糖,這件事看似精巧,卻藏著一個三十多億年沒被修好的破綻:負責固碳的核心酵素 Rubisco(ribulose-1,5-bisphosphate carboxylase/oxygenase)會「認錯受質」。它應該把 CO$_2$ 接到五碳的 RuBP 上,卻也會把 O$_2$ 接上去——這就是光呼吸(photorespiration)的源頭。理解光呼吸的代價,以及 C4、CAM 兩條獨立演化出來的「碳濃縮泵」如何規避它,是理解陸生植物地理分布與作物產量上限的關鍵。
Rubisco 的雙重活性可用競爭性受質的動力學描述。羧化(carboxylation)與氧化(oxygenation)共用同一活性位,兩者的相對速率取決於各自的催化效率與當地氣體濃度:
$$\frac{v_c}{v_o} = \frac{V_{c,max}/K_c}{V_{o,max}/K_o}\cdot\frac{[\mathrm{CO_2}]}{[\mathrm{O_2}]} = S_{c/o}\cdot\frac{[\mathrm{CO_2}]}{[\mathrm{O_2}]}$$
其中 $S_{c/o}$ 稱為「專一性因子(specificity factor)」,把 $V_{max}$ 與 Michaelis 常數 $K_m$ 整合成單一指標。C3 植物的 Rubisco $S_{c/o}$ 約落在 80–100(以 25°C 計)。值得注意的是:溫度升高時 CO$_2$ 在水中的溶解度下降得比 O$_2$ 快,且 Rubisco 的 $S_{c/o}$ 本身隨溫度下降——兩者疊加,使得高溫環境下氧化反應比例急遽上升。這正是光呼吸成為熱帶與乾旱植物演化壓力的物理根源。
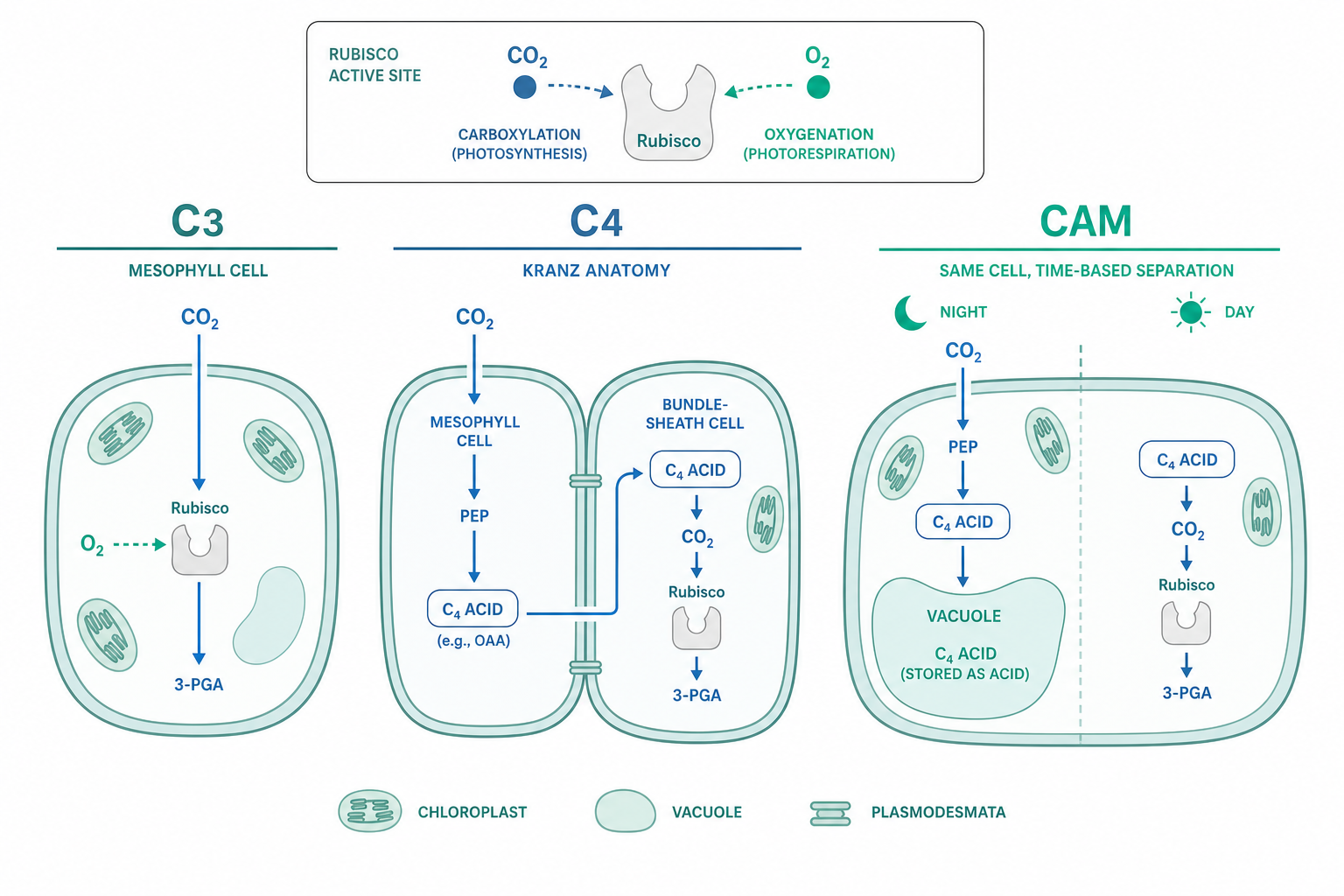
光呼吸途徑的能量帳:為什麼它是「淨損失」
當 Rubisco 氧化 RuBP,產物是一分子 3-磷酸甘油酸(3-PGA,可進 Calvin 循環)與一分子 2-磷酸乙醇酸(2-phosphoglycolate, 2-PG)。後者對細胞有毒——它會抑制磷酸丙糖異構酶與 Calvin 循環本身。植物因此演化出 C2 光呼吸途徑(photorespiratory carbon oxidation cycle),橫跨葉綠體、過氧化體(peroxisome)與粒線體三個胞器,把 2-PG 回收,但代價高昂:每處理兩分子 2-PG(即兩次氧化事件),會在粒線體經甘胺酸脫羧釋放出一分子已固定的 CO$_2$,並消耗 ATP 與還原力來再生。
把帳算清楚:C3 植物在 25°C、現今大氣(約 420 ppm CO$_2$、21% O$_2$)下,$v_o/v_c$ 約 0.25–0.3,意味著每固定約 3–4 個碳就要因光呼吸吐回 1 個碳。淨碳同化速率 $A$ 可近似寫成:
$$A = v_c\left(1 - \frac{0.5\,v_o}{v_c}\right) - R_d = v_c\left(1 - \frac{\Gamma_*}{C_i}\right) - R_d$$
其中 $\Gamma_*$ 是「CO$_2$ 補償點(無暗呼吸時)」,$C_i$ 是葉肉內 CO$_2$ 分壓。$\Gamma_*$ 在 C3 植物約 40–50 µbar 並隨溫度上升而升高;當 $C_i$ 逼近 $\Gamma_*$,淨光合趨近於零。這條 Farquhar–von Caemmerer–Berry(FvCB)模型方程式是當代植物生理量化光合的基石。
定量小範例:設某 C3 葉片 $v_c = 30\ \mu\text{mol·m}^{-2}\text{·s}^{-1}$,$v_o/v_c = 0.28$。光呼吸釋出 CO$_2$ 速率 $= 0.5\times v_o = 0.5\times(0.28\times30) = 4.2\ \mu\text{mol·m}^{-2}\text{·s}^{-1}$。總羧化貢獻 30 中被光呼吸抵銷 4.2,再扣暗呼吸 $R_d\approx1.5$,淨同化 $A\approx30-4.2-1.5=24.3\ \mu\text{mol·m}^{-2}\text{·s}^{-1}$。光呼吸單獨吃掉了約 14% 的毛固碳——在 35°C 高溫下此比例可逼近 30–40%。
C4 的空間隔離:把 CO$_2$ 泵到 Rubisco 面前
C4 植物(玉米、甘蔗、高粱、許多熱帶禾草)的解法是「空間區隔 + 生化泵」。它演化出 Kranz 解剖(花環狀構造):葉肉細胞與維管束鞘細胞(bundle sheath)兩種細胞分工。葉肉細胞中由 PEP 羧化酶(PEPC)先把 HCO$_3^-$ 固定成四碳的草醋酸——關鍵在於 PEPC 的受質是碳酸氫根而非 CO$_2$,完全不與 O$_2$ 競爭,且其對受質親和力極高($K_m$ 對 HCO$_3^-$ 約 5–30 µM)。
四碳酸(蘋果酸或天門冬胺酸)擴散進維管束鞘細胞後脫羧,在這個相對封閉的腔室裡把 CO$_2$ 濃度推高到大氣的 10–20 倍(可達 1000–2000 ppm)。此時 Rubisco 周遭的 $[\mathrm{CO_2}]/[\mathrm{O_2}]$ 比值被人為拉高,依前述競爭動力學,$v_o$ 被壓到幾乎可忽略——C4 植物的 $\Gamma_*$ 因此低到約 5–10 µbar。代價是每固定一個 CO$_2$ 多花 2 個 ATP(用於 PEP 再生)。在高溫強光下這筆額外能量投資划算;在低溫弱光下則 C3 反而較省,這解釋了 C4 禾草為何主宰熱帶草原而非寒帶。C4 光合至少獨立演化了 60 次以上,是趨同演化最壯觀的案例之一。
CAM 的時間隔離:把白天與黑夜拆開
CAM(景天酸代謝,Crassulacean Acid Metabolism)走的是「時間區隔」路線。仙人掌、鳳梨、龍舌蘭等乾旱植物在夜間開氣孔,用 PEPC 固碳並把蘋果酸大量儲存在液泡(使夜間細胞質明顯酸化);白天緊閉氣孔以減少蒸散失水,再把蘋果酸脫羧釋放 CO$_2$ 供 Rubisco 與 Calvin 循環使用。如此一來,光合在密閉氣孔的白天進行,CO$_2$ 內部濃度照樣被拉高、光呼吸被抑制,同時水分利用效率(WUE)可達 C3 植物的 3–6 倍。
C4 與 CAM 用的是同一套生化邏輯(PEPC 預固碳 + 脫羧釋放),差別只在隔離維度:C4 隔在空間,CAM 隔在時間。許多 CAM 植物還能依水分狀況在 C3 與 CAM 模式間切換(兼性 CAM),展現代謝可塑性。
深入探討(研究所視角)
Rubisco 的演化天花板與結構約束。為什麼演化沒有直接「修好」Rubisco?結構生物學給出機制性答案:羧化與氧化共用同一個由 Mg$^{2+}$ 配位、經胺甲醯化(carbamylation)活化的活性位,過渡態的幾何高度相似。Tcherkez 等人提出的「近乎完美(nearly perfect)」假說指出,$S_{c/o}$ 與羧化速率 $k_{cat}^c$ 之間存在強負相關的權衡(trade-off):要提高專一性,就得讓過渡態結合更緊、釋放更慢,催化轉換數因此下降。換言之,現存 Rubisco 已被推到此權衡曲線的 Pareto 前緣,這是熱力學與量子化學層次的硬約束,而非單純的「演化還沒來得及」。低溫紅藻 Griffithsia 的 Rubisco 擁有已知最高 $S_{c/o}$(約 167),是合成生物學移植改造的熱門模板,但其 $k_{cat}^c$ 也相應偏低,再次印證權衡的存在。
結構與組裝的瓶頸。高等植物 Rubisco 為 L8S8 全酶(八大、八小次單元),其折疊與組裝依賴特定分子伴侶(RbcX、Raf1、Raf2、BSD2 等)。長年無法在大腸桿菌中重組高等植物 Rubisco,直到近年釐清這套組裝因子的協同機制後才突破——這是把高效藍綠菌 Rubisco 或羧化體(carboxysome)的 CO$_2$ 濃縮機制移植進 C3 作物(如「C4 水稻計畫」與 carboxysome 工程)的前置技術。冷凍電鏡近年解析了藍綠菌 α-carboxysome 的殼蛋白幾何與內部 Rubisco 堆疊排列,揭示其如何形成擴散屏障把 CO$_2$ 局部濃縮,為人工碳濃縮構件提供藍圖。
單細胞 C4 與系統生物學重構。教科書說 C4 需要 Kranz 雙細胞解剖,但 Bienertia 與 Suaeda aralocaspica 等鹽生植物證明,單一細胞內可藉由葉綠體二態分化(dimorphic chloroplasts)與胞內擴散梯度,在同一個細胞質中完成 C4 循環——這顛覆了「C4 必須有 Kranz」的舊典範。系統生物學層面,比較轉錄體與單細胞 RNA-seq 顯示,從 C3 到 C4 的轉變多半不是發明新基因,而是把既有基因(PEPC、NADP-malic enzyme、carbonic anhydrase)的表現量、細胞特異性與啟動子調控重新佈線(rewiring),這使「人工誘導 C3 作物部分 C4 化」在理論上可行。代謝通量分析($^{13}$C-MFA)配合 FvCB 模型,已能在不同光溫梯度下量化 C4 兩室間 CO$_2$ 滲漏率(leakiness, $\phi$,典型約 0.2–0.3),這個參數直接決定 C4 額外 ATP 投資的能量效率,是評估工程化策略成敗的核心量化指標。
演化基因體學的近期視角。系統發育重建顯示 C4 的反覆獨立起源往往經過 C3–C4 中間型過渡階段(如 Flaveria 屬),這些中間型先把甘胺酸脫羧酶(GDC)侷限到維管束鞘細胞,形成「光呼吸 CO$_2$ 泵(C2 photosynthesis)」雛形,再逐步疊加完整 C4 生化——為「複雜性狀如何分步演化」提供了活生生的連續譜系,也是當前作物工程試圖複製的演化路徑圖。