
酵素動力學:Michaelis–Menten 與抑制的量化解析
從穩態假設到別構協同,看懂 Km、kcat 與四種抑制模式的分子指紋
為什麼一個酵素「跑不快也壓不住」?
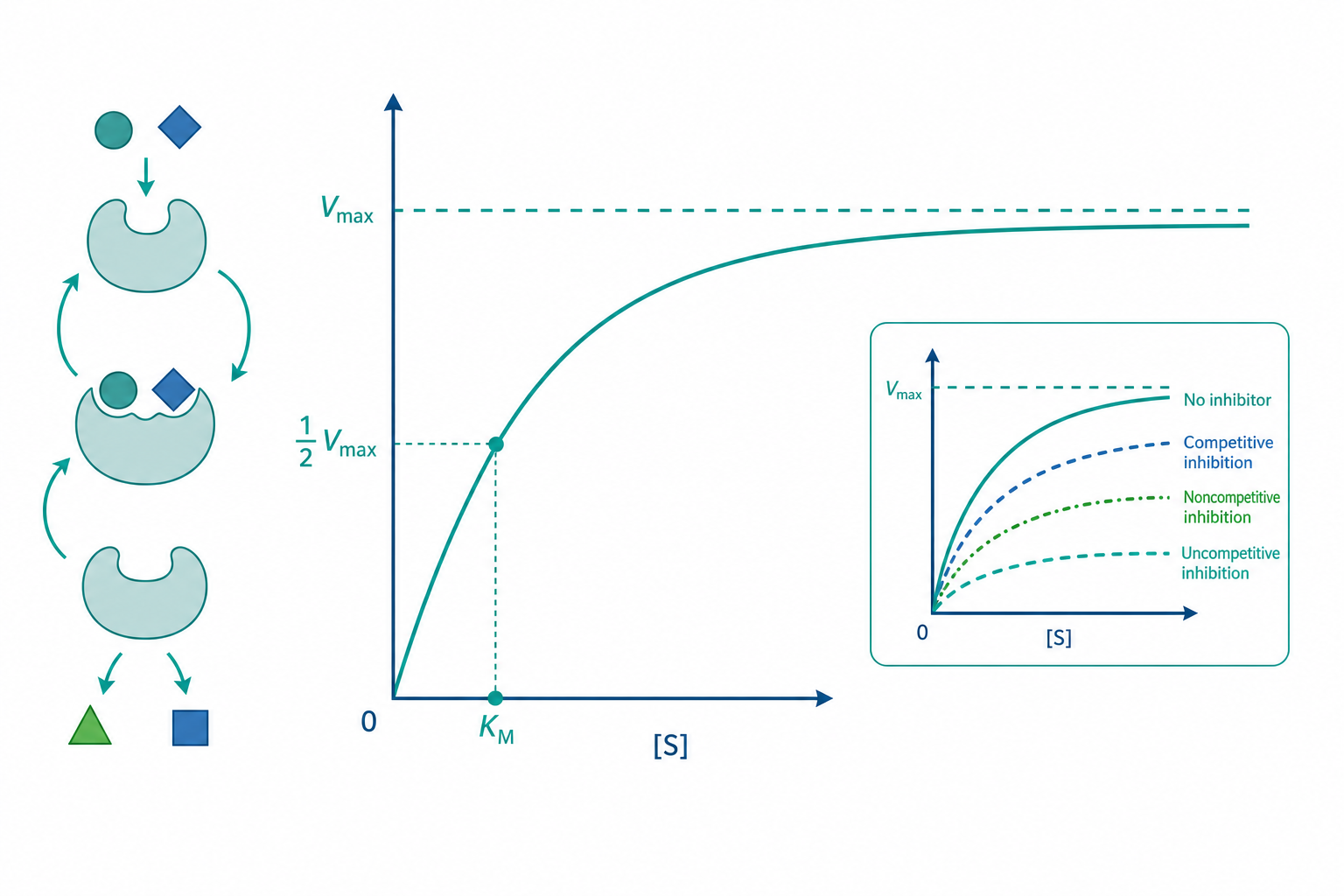
直覺上,我們以為酵素越多受質就反應越快——加倍受質,速度就加倍。但真實的酵素並非如此。當受質濃度爬升到某個程度後,反應速度會「撞牆」趨於平頂,再怎麼加受質也擠不出更多產物。這個飽和現象,正是酵素催化的核心特徵:酵素的活性位點數量有限,催化是一個「結合—轉化—釋放」的循環,而非無上限的線性放大。要把這件事說清楚,我們需要的不只是定性描述,而是一個能寫成方程式、能從實驗數據反推參數、能預測抑制劑如何改寫整條曲線的量化框架。這就是 Michaelis–Menten 動力學(米氏動力學)登場的地方。
本篇假設你已具備「酵素是生物催化劑、會降低活化能」的基礎,因此我們直接從機制與量化模型切入,討論米氏方程的推導假設、$K_m$ 與 $k_{cat}$ 的真正物理意義,以及四種抑制模式如何在 Lineweaver–Burk 圖上留下截然不同的指紋。
米氏方程的推導與穩態假設
考慮最簡單的單受質反應機制:
$$E + S \xrightleftharpoons[k_{-1}]{k_1} ES \xrightarrow{k_{cat}} E + P$$
酵素 $E$ 與受質 $S$ 可逆地形成複合物 $ES$,後者再以速率常數 $k_{cat}$(又稱轉換數,turnover number)不可逆地釋出產物 $P$。1925 年 Briggs 與 Haldane 提出的穩態假設(steady-state approximation)指出:反應啟動後極短時間內,$ES$ 的生成速率與分解速率達到平衡,使 $\frac{d[ES]}{dt} \approx 0$。
由此可解出初速度(initial velocity):
$$v_0 = \frac{V_{max}[S]}{K_m + [S]}$$
其中 $V_{max} = k_{cat}[E]_T$ 是酵素被受質完全飽和時的最大速度,而米氏常數定義為:
$$K_m = \frac{k_{-1} + k_{cat}}{k_1}$$
這條雙曲線方程精準地描述了前述的飽和行為。當 $[S] \ll K_m$ 時,$v_0 \approx \frac{V_{max}}{K_m}[S]$,呈一級反應、速度與受質濃度成正比;當 $[S] \gg K_m$ 時,$v_0 \approx V_{max}$,呈零級反應、速度與受質濃度無關。
$K_m$ 的物理意義常被誤解。它的數值等於使反應達到 $\frac{1}{2}V_{max}$ 時的受質濃度,是一個具有濃度單位的常數。$K_m$ 並不直接等於解離常數 $K_d = k_{-1}/k_1$;只有在 $k_{cat} \ll k_{-1}$(即催化遠慢於複合物解離)這個特例下,$K_m$ 才近似於 $K_d$,此時 $K_m$ 才可解讀為親和力的倒數指標。
催化效率:$k_{cat}/K_m$ 與擴散極限
衡量酵素「好不好用」不能只看 $k_{cat}$ 或只看 $K_m$。真正具生理意義的指標是特異性常數(specificity constant)$k_{cat}/K_m$,它是低受質濃度下的二級速率常數,反映酵素在「受質稀薄、彼此競爭」的細胞真實環境中對某受質的整體催化偏好。
$k_{cat}/K_m$ 有一個物理上限:它不可能超過 $E$ 與 $S$ 在溶液中相遇的擴散速率,約 $10^8 \sim 10^9 \ \mathrm{M^{-1}s^{-1}}$。已知接近此「擴散極限」的酵素如三醣磷酸異構酶(triosephosphate isomerase)、過氧化氫酶(catalase)與乙醯膽鹼酯酶,被稱為催化完美(catalytically perfect)的酵素——它們的速率瓶頸已不在化學步驟,而在受質擴散送達的速度本身。
四種抑制模式的量化指紋
抑制劑透過不同方式插手反應,可由它們對 $V_{max}$ 與 $K_m$ 的影響來分類。引入抑制因子 $\alpha = 1 + \frac{[I]}{K_i}$(作用於游離酵素)與 $\alpha' = 1 + \frac{[I]}{K_i'}$(作用於 $ES$ 複合物),可寫出統一形式:
$$v_0 = \frac{V_{max}[S]}{\alpha K_m + \alpha'[S]}$$
- 競爭型抑制(competitive):抑制劑與受質競爭同一活性位點,僅作用於游離 $E$($\alpha > 1, \alpha' = 1$)。表觀 $K_m$ 上升,$V_{max}$ 不變(足量受質可把抑制劑排擠掉)。
- 非競爭型抑制(noncompetitive):抑制劑結合於別處,對 $E$ 與 $ES$ 親和力相同($\alpha = \alpha' > 1$)。表觀 $V_{max}$ 下降,$K_m$ 不變。
- 反競爭型抑制(uncompetitive):抑制劑只結合 $ES$($\alpha = 1, \alpha' > 1$)。$V_{max}$ 與 $K_m$ 同比例下降,$\frac{V_{max}}{K_m}$ 不變。
- 混合型抑制(mixed):$\alpha \neq \alpha'$ 且皆大於 1,$V_{max}$ 必降,$K_m$ 視兩者大小而升或降。
在 Lineweaver–Burk 雙倒數圖($\frac{1}{v_0}$ 對 $\frac{1}{[S]}$)上,這些模式各有特徵:競爭型的直線族交於 $y$ 軸同一點($V_{max}$ 不變);非競爭型交於 $x$ 軸同一點($K_m$ 不變);反競爭型則呈一組平行線。
一個定量小範例
假設某酵素 $V_{max} = 100 \ \mu\mathrm{M/min}$、$K_m = 2 \ \mathrm{mM}$。
問一:當 $[S] = 2 \ \mathrm{mM}$ 時的初速度? 代入米氏方程:$v_0 = \frac{100 \times 2}{2 + 2} = 50 \ \mu\mathrm{M/min}$,恰為 $\frac{1}{2}V_{max}$——驗證了 $K_m$ 的定義。
問二:加入競爭型抑制劑 $[I] = 4 \ \mathrm{mM}$、$K_i = 2 \ \mathrm{mM}$,在 $[S] = 2 \ \mathrm{mM}$ 下速度為何? 先算 $\alpha = 1 + \frac{4}{2} = 3$。表觀 $K_m^{app} = \alpha K_m = 6 \ \mathrm{mM}$。 $$v_0 = \frac{100 \times 2}{6 + 2} = 25 \ \mu\mathrm{M/min}$$ 速度從 50 降到 25。注意:若把 $[S]$ 拉到極高,$v_0$ 仍會回升趨近 100——這正是競爭型抑制「可被受質克服」的標誌,與非競爭型的天花板下降形成對照。
別動力學:當曲線不再是雙曲線
米氏框架假設酵素只有單一獨立活性位點。但許多代謝調控的關鍵酵素是多次單元的別構酶(allosteric enzyme),其速度曲線呈 S 形(sigmoidal)而非雙曲線,反映次單元間的協同結合(cooperativity)。此時改用 Hill 方程描述:
$$\theta = \frac{[S]^n}{K^n + [S]^n}$$
其中 Hill 係數 $n > 1$ 代表正協同。經典範例是血紅素的氧解離曲線(雖屬運輸蛋白而非酵素,但協同數學相同),以及糖解途徑的限速酵素磷酸果糖激酶(PFK),後者受 ATP 與 AMP 的別構回饋調控,是細胞能量感測的核心節點。
深入探討(研究所視角)
進入研究所層次,米氏動力學從一個「擬合工具」轉變為通往酵素機制與演化的入口。以下幾個方向是主體刻意未觸及的前沿。
過渡態理論與催化的真正來源。 酵素的催化威力不在於穩定受質,而在於選擇性地穩定過渡態(transition state)。Linus Pauling 早在 1948 年即提出此洞見:若酵素對過渡態的親和力遠高於對基態受質的親和力,則 $k_{cat}/K_m$ 的提升幅度可由過渡態結合自由能差直接預測。這也解釋了過渡態類似物(transition-state analogs)為何是極強的抑制劑——它們模擬過渡態幾何,結合常數可達奈莫耳甚至皮莫耳級。催化抗體(abzymes)的設計正是基於此原理:用過渡態類似物作 hapten 誘導抗體,使其獲得催化活性。
結構生物學與時間解析晶體學。 X 射線晶體學、低溫電子顯微鏡(cryo-EM)與時間解析串列飛秒晶體學(time-resolved serial femtosecond crystallography, 使用 X 射線自由電子雷射 XFEL)已能「拍下」催化循環中的瞬態構象,捕捉到誘導契合(induced fit)與構象選擇(conformational selection)兩種模型的實際軌跡。這推翻了酵素是剛性「鎖鑰」的舊圖像,揭示活性位點是動態的,催化伴隨毫秒至皮秒尺度的構象運動。
動力學同位素效應與量子穿隧。 將受質上的氫換成氘(²H)或氚(³H),可量測一級動力學同位素效應(KIE)。異常巨大的 KIE(遠超半古典上限約 7)是氫原子量子穿隧(quantum tunnelling)的指紋。在醇脫氫酶等系統中,已有強力證據顯示氫轉移部分透過穿隧進行,且酵素的構象動態會調制能障寬度以促進穿隧——這把酵素催化推進到量子生物學的領域。
單分子酵素學與動態異質性。 傳統米氏實驗量測的是數以億計酵素分子的系綜平均。單分子螢光技術(如基於 FRET 或產物螢光的方法)揭示了動態異質性(dynamic disorder):同一個酵素分子的 $k_{cat}$ 會隨時間在不同構象態間漲落,且這種記憶可持續數秒。系綜層次的米氏雙曲線,竟可在單分子隨機軌跡下嚴格重現——這項理論成果(Xie 等人的工作)深化了我們對「速率常數」本質的理解。
系統生物學與代謝控制分析。 在整條代謝途徑的尺度上,單一酵素的 $K_m$ 與 $V_{max}$ 不足以預測通量(flux)的控制權落在何處。代謝控制分析(Metabolic Control Analysis, MCA)以「通量控制係數」量化每個酵素對整體途徑通量的貢獻,並由連加定理約束(所有係數總和為 1)。其核心結論顛覆直覺:途徑中往往沒有單一「限速步驟」,控制權是分散且依條件變動的。這套框架結合單細胞體學與動力學模型,是當代代謝工程與系統生物學設計合成途徑的理論基石。