
適應性免疫的分子機制:MHC 與 T/B 細胞的量化邏輯
從 V(D)J 重組、動力學校對到生發中心的親和力成熟
從「亂槍打鳥」到「狙擊手」:適應性免疫的分子邏輯
固有免疫像守門的保全,靠少數預設的模式辨識受體攔截病原;適應性免疫則像可以為每一個敵人「量身打造武器」的軍火庫。它的核心難題只有一個:宿主基因組只有約兩萬個蛋白質編碼基因,卻必須準備出足以辨識 $10^{11}$ 種以上抗原表位的受體庫。本文要回答的,正是這套系統如何在分子層次用有限的基因生成近乎無限的辨識能力,又如何用熱力學與動力學的語言,把「辨識強弱」轉換成可量化的細胞命運決策。
換句話說,適應性免疫不只是「記住敵人」這麼直覺的故事,而是一個由親和力閾值、結合動力學與群體選擇共同驅動的量化系統。
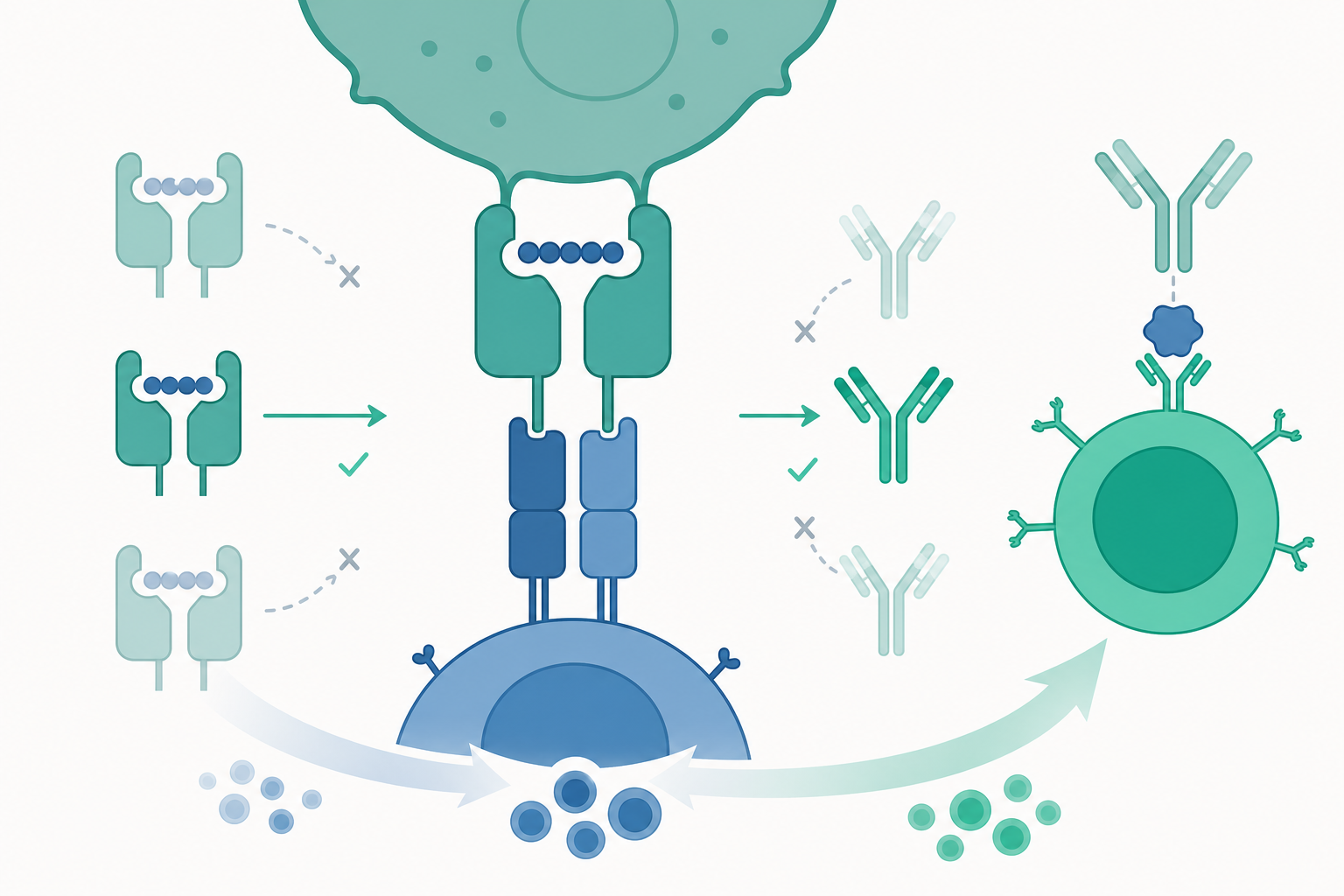
V(D)J 重組:用組合數學生成受體多樣性
B 細胞受體(BCR/抗體)與 T 細胞受體(TCR)的多樣性,來自體細胞層次的基因重排。重鏈可變區由 V、D、J 三段基因片段隨機拼接,輕鏈由 V、J 兩段拼接,由 RAG1/RAG2 重組酶識別重組訊號序列(RSS)後切割、再連接。
若僅計算組合多樣性,以人類重鏈約 $V=40$、$D=25$、$J=6$ 片段、輕鏈約 $V=70$、$J=9$ 估計,純片段組合的數量級為:
$$N_{\text{combinatorial}} = (40 \times 25 \times 6) \times (70 \times 9) \approx 3.8 \times 10^{6}$$
但真正讓多樣性爆炸的是接合區多樣性(junctional diversity):TdT 酶在接點隨機加入 N 核苷酸、外切酶削去 P 核苷酸,使 CDR3 區序列幾乎不重複。算上接合多樣性與重輕鏈配對,理論受體庫可達 $10^{13}$ 以上,遠超過 B 細胞實際數目(約 $10^{11}$)。這正是「組合爆炸」如何用幾十個片段換來天文數字的核心。
MHC 限制與胜肽呈現的熱力學
T 細胞不能直接辨識游離抗原,必須辨識「MHC 分子凹槽中呈現的胜肽片段」,這就是 MHC 限制(MHC restriction)。MHC I 類(呈現給 CD8⁺ T 細胞)通常結合 8–10 個胺基酸的胜肽,凹槽兩端封閉;MHC II 類(呈現給 CD4⁺ T 細胞)凹槽兩端開放,可容納 13–25 個殘基。
胜肽與 MHC 的結合可視為平衡反應,其穩定性決定了表位能否被有效呈現。以解離常數 $K_d$ 描述:
$$\Delta G^{\circ} = -RT \ln\!\left(\frac{1}{K_d}\right) = RT \ln K_d$$
一個高親和力表位的 $K_d$ 可低至奈莫耳級。取 $K_d = 10^{-8}\,\text{M}$、$T = 310\,\text{K}$、$R = 8.314\,\text{J·mol}^{-1}\text{K}^{-1}$:
$$\Delta G^{\circ} = (8.314)(310)\ln(10^{-8}) \approx 2577 \times (-18.42) \approx -47.5\,\text{kJ·mol}^{-1}$$
負值越大代表結合越穩定、胜肽停留在表面越久,T 細胞越有機會掃描到它。MHC 的另一關鍵是多基因性與多型性:人類 HLA-A/B/C、HLA-DR/DQ/DP 是已知最具多型性的基因座,等位基因數以萬計。在族群層次,這種多型性可用 Hardy–Weinberg 平衡描述,若某 HLA 等位基因頻率為 $p$,異型合子頻率為 $2p(1-p)$,異型合子能呈現更廣的胜肽譜——這被認為是 HLA 維持高度多型性的演化驅動力(異型合子優勢)。
TCR 觸發的動力學校對:kinetic proofreading
TCR 如何在親和力差異極小的情況下,區分自我與非自我胜肽?關鍵不是單純的「結合強度」,而是結合的停留時間(dwell time)。McKeithan 提出的動力學校對(kinetic proofreading)模型指出:TCR 與 pMHC 結合後,必須經過一連串磷酸化中間步驟(由 Lck、ZAP-70 介導)才能送出活化訊號;若在抵達終點前 pMHC 提早解離,整串磷酸化會重置。
設每一步前進速率為 $k_p$、解離速率為 $k_{\text{off}}$,則完成 $N$ 步校對的機率為:
$$P_{\text{signal}} = \left(\frac{k_p}{k_p + k_{\text{off}}}\right)^{N}$$
由於停留時間 $\tau = 1/k_{\text{off}}$,此式對 $k_{\text{off}}$ 高度非線性敏感:解離稍快($\tau$ 稍短),$P_{\text{signal}}$ 便以指數方式崩潰。這解釋了為何 TCR 能把奈秒到秒級的停留時間差,放大成「活化/不活化」的二元決策——它本質上是一個生化層次的低通濾波與訊號放大器。
B 細胞的親和力成熟:達爾文式的微演化
抗原進入淋巴結後,B 細胞在生發中心(germinal center)經歷「暗區增殖/突變、亮區選擇」的循環。AID(活化誘導胞苷脫氨酶)在可變區引入體細胞超突變,突變率高達每代每鹼基對 $10^{-3}$,比一般基因組高約百萬倍。
這是一個典型的選擇方程式問題:帶有較高親和力 BCR 的 B 細胞,能從濾泡樹突細胞奪取更多抗原、向 $T_{FH}$ 細胞呈現更多 pMHC,獲得更強的存活與增殖訊號。經數輪循環,群體平均親和力可提升 $10$ 到 $1000$ 倍,$K_d$ 從微莫耳級降到皮莫耳級。
定量小範例:假設初始 B 細胞群中高親和力選殖佔比 $x_0 = 0.01$,其相對適存度(每輪存活增殖優勢)為 $w = 1.5$。在簡化的離散選擇模型中,$n$ 輪後高親和力選殖佔比約為:
$$x_n = \frac{x_0\, w^{n}}{x_0\, w^{n} + (1 - x_0)}$$
代入 $n = 10$:$w^{10} = 1.5^{10} \approx 57.7$,分子 $= 0.01 \times 57.7 = 0.577$,分母 $= 0.577 + 0.99 = 1.567$,故 $x_{10} \approx 0.368$。短短十輪選擇,高親和力選殖就從 1% 上升到約 37%——這就是生發中心如何在數週內把抗體「磨利」的數學直覺。
訊號整合與類別轉換
完整活化需「兩個訊號」:T 細胞需要 TCR–pMHC(訊號一)加上 CD28–B7 共刺激(訊號二),缺一則進入無能(anergy)狀態,這是外周耐受的重要機制。B 細胞則在 $T_{FH}$ 提供的 CD40L 與細胞激素環境下進行類別轉換重組(CSR),把 IgM 換成 IgG/IgA/IgE,同樣由 AID 引發重鏈恆定區的 DNA 雙股斷裂與重接。最終,記憶 B 細胞與長壽漿細胞固化下來,讓再次感染時的反應在數天內就壓制病原——這便是疫苗有效的分子根據。
深入探討(研究所視角)
結構生物學:TCR 觸發的力學與構象模型。 動力學校對雖能解釋活化的非線性,但無法完全說明「催化力鍵(catch bond)」現象——單分子力譜(光鑷、AFM、生物膜力探針)顯示某些 agonist pMHC 在受到 pN 級拉力時,TCR–pMHC 鍵的壽命不減反增,而 antagonist 則形成滑鍵(slip bond)。這支持了 TCR 的力學感測(mechanosensing)假說:T 細胞遷移與膜起伏施加的剪切力,是區分自我與非自我的物理過濾器。冷凍電鏡近年解出 TCR–CD3 完整複合體的跨膜排列,顯示 CD3 二聚體如何包裹 TCR 並把胞外結合事件傳遞為胞內 ITAM 構象變化,為「allosteric vs. mechanical」之爭提供了結構基礎。
單細胞與體學:解構受體庫與選殖動態。 單細胞 TCR/BCR 定序(scRNA-seq + VDJ-seq)讓我們能在同一細胞同時讀出受體序列、轉錄體與表觀狀態,重建生發中心選殖的系統發生樹(lineage tree)。配合質譜流式(CyTOF)與空間轉錄體(如 MERFISH、Visium),研究者能在淋巴結原位定位暗區/亮區的選殖遷移。免疫受體庫定序(AIRR-seq)則把整個受體庫當成可統計的群體,用 Shannon 多樣性、clonality、公共選殖(public clones)等指標量化個體間共享程度,並追蹤疫苗或感染後的選殖擴張軌跡。
系統生物學模型:閾值、交叉反應與耐受。 「擾動–親和力」(perturbation–affinity)與 TCR 交叉反應的定量研究指出,單一 TCR 平均可辨識 $10^{4}$–$10^{6}$ 種不同 pMHC,受體庫的辨識覆蓋因此來自高度簡併(degeneracy)而非一對一專一。胸腺的正負選擇可建模為對自我 pMHC 親和力的「窗口函數」:親和力過低者凋亡(無法存活)、過高者被刪除或轉為 Treg,僅落在中間窗口者輸出。這種閾值選擇與 Treg 的外周抑制,共同構成可用微分方程描述的耐受網路,其穩態分析有助於理解自體免疫的失衡點。
近期機制進展。 在治療層面,這些分子原理已被工程化:嵌合抗原受體(CAR)把抗體的單鏈可變片段(scFv)接上 CD3ζ 與共刺激域,繞過 MHC 限制直接辨識表面抗原;雙特異性 T 細胞銜接器(BiTE)則人工拉近 T 細胞與腫瘤。而結構為本的免疫原設計(如以穩定構象的醣蛋白三聚體引導生發中心朝特定 broadly neutralizing 路徑成熟)正把「設計受體庫的演化軌跡」變為可操作的工程問題。這些都共享同一個量化骨架:親和力、停留時間與選擇壓力,三者決定了適應性免疫的每一個分子決策。