
細胞的海關:磷脂雙層如何控制萬物進出
從擴散、滲透到鈉鉀幫浦,看一層薄膜如何撐起整個生命
細胞的「海關」:為什麼一層膜這麼重要?
想像你住的城市四周圍著一道城牆,城牆上有許多閘門。有些東西可以自由進出,有些得排隊查驗,有些則要花錢請人扛進來。這道城牆與閘門系統,決定了城市的安全與運作——對一顆細胞來說,扮演這個角色的就是細胞膜。
細胞膜是包裹在細胞最外層的一層薄膜,厚度只有約 7 到 10 奈米(一奈米是十億分之一公尺)。別小看它,沒有這層膜,細胞就無法把「自己」和「外面」分開,更無法維持內部穩定的環境。生命之所以能存在,第一步就是「圍出一個可以控制進出的空間」。
而控制進出,正是這篇文章的主角:物質運輸。
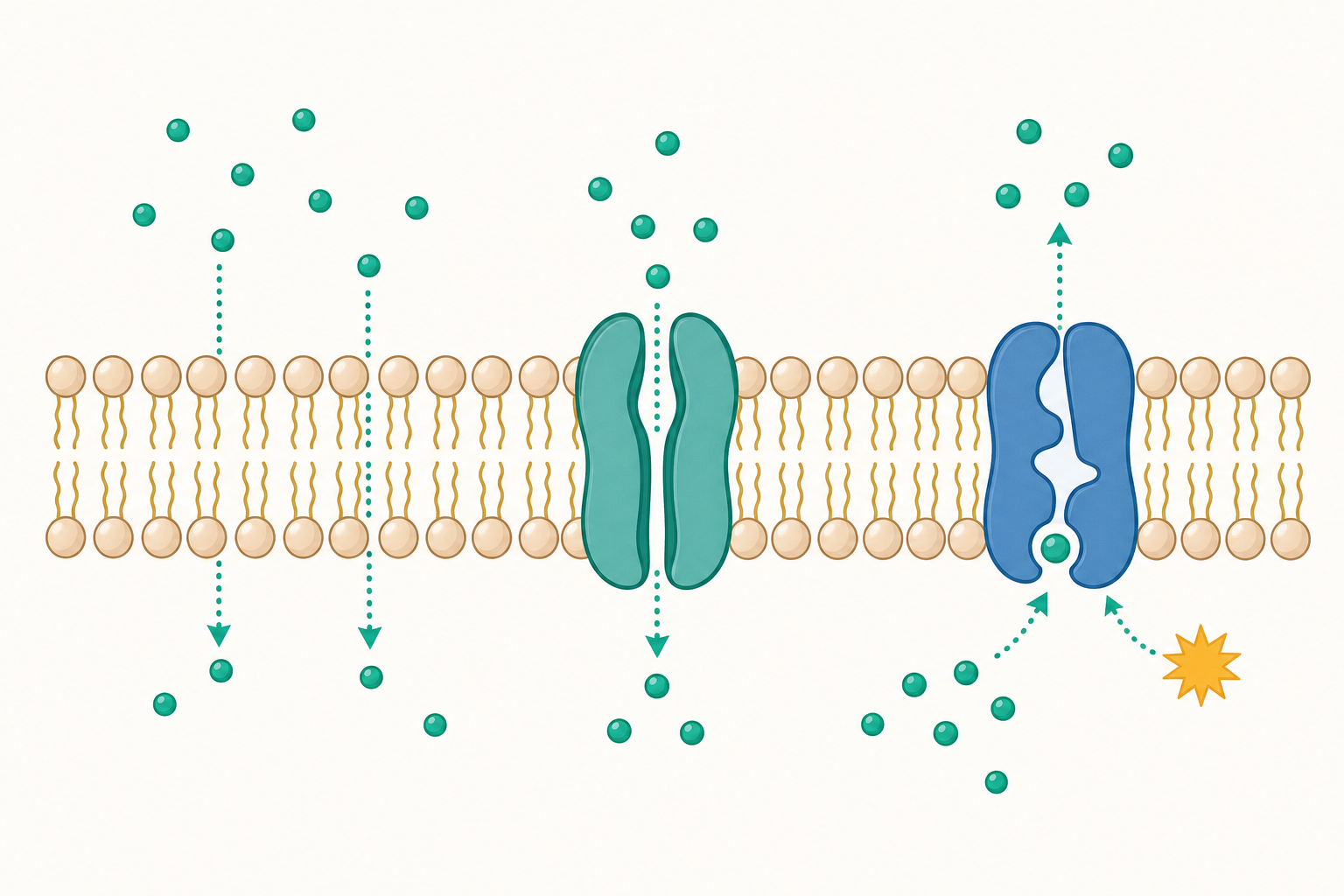
磷脂雙層:一道又「怕水」又「親水」的牆
細胞膜的主要建材是磷脂(phospholipid)。每一個磷脂分子都有兩個性格相反的部分:
- 頭部:帶有磷酸基,喜歡水,稱為「親水性」(hydrophilic)。
- 尾部:兩條脂肪酸長鏈,討厭水,稱為「疏水性」(hydrophobic)。
當大量磷脂分子泡在水裡時,它們會自動排成兩層:所有的「怕水尾巴」躲在中間互相靠攏,所有的「親水頭部」朝向內外兩側的水。這就形成了著名的磷脂雙層(phospholipid bilayer)。
這個結構非常聰明。中間那層油性的尾巴,像一道防水夾層,讓水溶性的離子(如鈉離子 $\text{Na}^+$、鉀離子 $\text{K}^+$)和大分子無法輕易穿過。但小分子、不帶電的氣體(如氧氣 $\text{O}_2$、二氧化碳 $\text{CO}_2$)卻能直接溶進去穿越。
膜上還鑲嵌著各種蛋白質,有的當「通道」,有的當「幫浦」,有的負責辨識訊號。科學家把這種「磷脂像流動的海、蛋白質像漂浮的船」的模型稱為流體鑲嵌模型(fluid mosaic model)。膜不是死板的牆,而是會流動、會變形的動態結構。
不花力氣的運輸:擴散與促進擴散
物質穿過細胞膜的方式,可以先用一個問題來分類:細胞要不要消耗能量?
不需要消耗能量、順著濃度「由高往低」流動的,叫做被動運輸(passive transport)。最基本的形式就是擴散(diffusion)。
把一滴墨水滴進清水,墨水會自己慢慢散開,直到整杯水均勻變色。這就是擴散:分子總是傾向從濃度高的地方移動到濃度低的地方,直到兩邊一樣多。氧氣進入細胞、二氧化碳排出細胞,靠的就是這種單純擴散。
但有些分子(如葡萄糖、離子)雖然也想順濃度差移動,卻因為帶電或太大而過不了那層油牆。這時就需要膜上的蛋白質「開門」幫忙,稱為促進擴散(facilitated diffusion)。注意:雖然有蛋白質參與,但方向仍是由高濃度往低濃度,細胞不需要花能量,所以仍屬被動運輸。
滲透:水的特別待遇
水分子的移動是被動運輸中特別重要的一種,專門有個名字叫滲透(osmosis):水會從「溶質濃度低」的一側,穿過半透膜,移向「溶質濃度高」的一側。
用一個生活對照來理解,把細胞放進三種不同的溶液:
| 外界溶液 | 溶質濃度比較 | 水的淨移動 | 細胞(以紅血球為例) |
|---|---|---|---|
| 低張溶液 | 外低內高 | 水往內流 | 膨脹,甚至脹破 |
| 等張溶液 | 內外相等 | 無淨移動 | 維持正常形狀 |
| 高張溶液 | 外高內低 | 水往外流 | 皺縮 |
這就是為什麼醃漬食物能防腐——高濃度的鹽或糖把微生物細胞裡的水「吸」出來,讓它脫水而無法生存。也是為什麼點滴注射要用「生理食鹽水」(與血液等張),否則紅血球會脹破或皺縮。
要花力氣的運輸:主動運輸與鈉鉀幫浦
如果細胞想把某種物質「逆著濃度差」搬運——也就是從少的一邊搬到多的一邊,就像把水往高處推——那就違反自然趨勢了,必須消耗能量。這叫做主動運輸(active transport)。
細胞的能量貨幣是 ATP(腺苷三磷酸)。主動運輸的經典例子是鈉鉀幫浦(sodium-potassium pump)。它每消耗一個 ATP,就把:
$$3\,\text{Na}^+ \;(\text{往細胞外}) \quad\text{與}\quad 2\,\text{K}^+ \;(\text{往細胞內})$$
逆著各自的濃度差運出與運入。神經細胞傳遞訊號、肌肉收縮,都仰賴這個幫浦維持細胞內外的離子差距。可以說,我們的每一個念頭、每一次心跳,背後都有無數個鈉鉀幫浦在默默耗能工作。
對於更大型的物質(如整顆病毒、大團蛋白質),細胞甚至會用膜把它「包起來」吞進去,稱為胞吞(endocytosis);反過來把物質包成小泡送出去,則稱為胞吐(exocytosis)。
小結:細胞如何當好一位「海關」
回到城牆的比喻,我們可以把三種運輸方式整理成一條清楚的判斷流程:
- 分子小、不帶電? → 直接穿過磷脂雙層(單純擴散)。
- 帶電或太大,但順濃度差? → 走蛋白質通道,不耗能(促進擴散)。
- 要逆濃度差? → 動用 ATP 幫浦(主動運輸)。
- 是水分子? → 順著溶質濃度差移向高張側(滲透)。
細胞膜不只是被動的圍牆,更是一位精明的海關:該放行的放行、該攔的攔、該花錢扛進來的也絕不手軟。正是這種對「進出」的精密控制,讓細胞得以維持穩定的內環境,撐起整個生命的運作。
深入探討(研究所視角)
入門段把運輸分成「耗能/不耗能」兩大類,但在分子層級,更精準的框架是電化學梯度(electrochemical gradient)。離子的跨膜流動同時受到兩股力量影響:濃度梯度(化學項)與膜電位(電學項)。這兩者合起來的能量差,可用以下關係描述:
$$\Delta G = RT \ln\frac{[C]_{in}}{[C]_{out}} + zF\Delta\psi$$
其中 $z$ 為離子電荷、$F$ 為法拉第常數、$\Delta\psi$ 為膜電位、$R$ 為氣體常數、$T$ 為絕對溫度。當 $\Delta G < 0$ 時離子自發流動,$\Delta G > 0$ 時則需外加能量。對單一帶電離子達到平衡時,這個式子可化簡為著名的能斯特方程(Nernst equation),用來計算神經細胞的平衡電位;進一步考慮多種離子的同時通透,則導出 GHK 方程(Goldman–Hodgkin–Katz equation),這是理解神經與肌肉靜息電位的理論基石。
主動運輸本身也可細分。鈉鉀幫浦這類直接水解 ATP 來驅動的,稱為初級主動運輸(primary active transport),屬於 P-type ATPase。而許多養分的吸收則靠次級主動運輸(secondary active transport):細胞先用初級幫浦建立鈉離子梯度,再讓鈉離子「順流而下」時順便把葡萄糖「逆流帶上」——這就是腸道與腎臟 SGLT 共同運輸蛋白(co-transporter)的工作原理。同向搬運稱 symport,反向稱 antiport,能量是「借來的」而非直接消耗 ATP。
水的運輸在 1990 年代被改寫。過去認為水只能緩慢滲透過磷脂,直到 Peter Agre 發現水通道蛋白(aquaporin)這一專門讓水快速通過的膜蛋白,並因此獲 2003 年諾貝爾化學獎(與研究離子通道的 Roderick MacKinnon 共享)。MacKinnon 利用 X 光晶體學解出鉀離子通道的原子結構,揭示通道如何以「選擇性過濾器」精準辨別 $\text{K}^+$ 與較小的 $\text{Na}^+$——這是結構生物學詮釋膜功能的里程碑。
當代研究更把細胞膜推向系統與工程層次。膜片鉗技術(patch clamp,Neher 與 Sakmann 於 1991 年獲諾貝爾獎)讓人能記錄單一通道開關的電流;冷凍電顯(cryo-EM)近年解析出大量運輸蛋白的高解析結構,揭露其構型變化的動態週期。在合成生物學中,研究者嘗試以人工脂質體與重組通道蛋白打造最小細胞,探討維持跨膜梯度所需的最少元件。膜運輸的異常也直接連結疾病:囊狀纖維化源於氯離子通道 CFTR 的突變,許多抗癌與抗藥性問題則與 ABC 運輸蛋白把藥物「主動泵出」細胞有關。從一層薄薄的磷脂,延伸到電生理、結構生物學乃至藥物設計——細胞膜始終是分子生命科學最豐富的舞台之一。