
內膜系統與蛋白質分泌途徑:從共轉譯轉位到囊泡運輸的分子物流
以動力學、自由能與品質管制量化解析真核細胞的分泌工廠
從一條多肽鏈說起:分泌途徑是一場精密的物流系統
想像細胞要把一個剛合成的酵素送出細胞外,這趟旅程不是「丟出去」這麼簡單。一條多肽鏈從核糖體誕生的那一刻起,就被打上標籤、層層檢核、逐站加工,最終封裝進囊泡送達目的地。這套貫穿內質網(ER)、高基氏體(Golgi)、囊泡與細胞膜的運輸網絡,就是內膜系統(endomembrane system)。它承擔了真核細胞約三分之一蛋白質體的成熟與運輸,是「分泌途徑(secretory pathway)」的物理載體。
進階地說,分泌途徑的核心問題不是「蛋白質會不會被送出去」,而是「在什麼速率、以什麼通透性、付出多少自由能成本」下完成精準分選。本文將從共轉譯轉位(co-translational translocation)的機制談起,逐步深入到囊泡運輸的動力學與品質管制的量化邏輯。
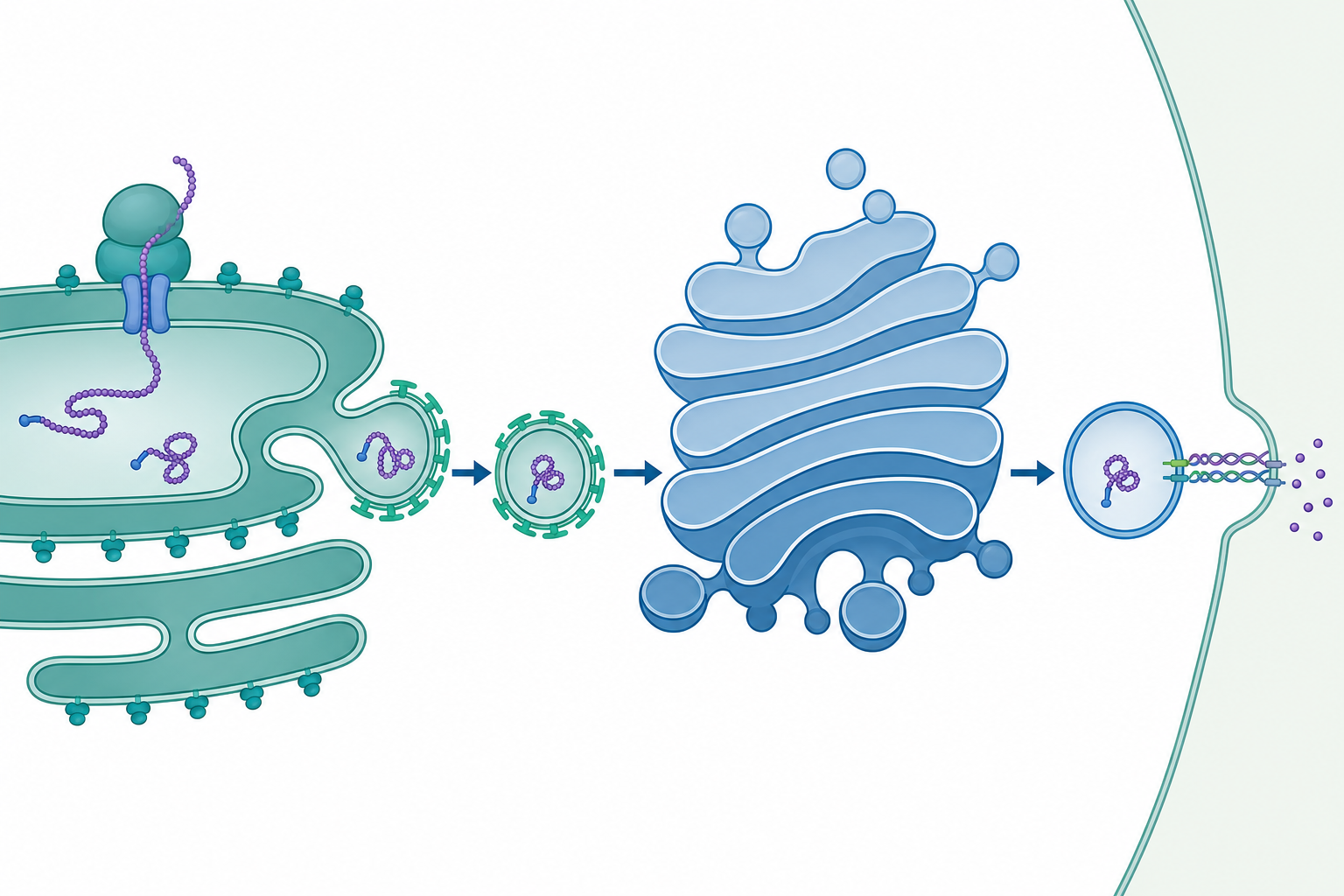
訊號識別與共轉譯轉位:第一道閘門
分泌蛋白與膜蛋白的 N 端通常帶有一段疏水的訊號胜肽(signal peptide)。當它從核糖體出口通道露出,訊號識別顆粒(SRP)便辨識並結合,暫停轉譯(translational arrest)。SRP 引導核糖體–新生鏈複合體靠泊到 ER 膜上的 SRP 受體,再交棒給 Sec61 轉位子(translocon)——一個漏斗狀的蛋白質通道。多肽鏈隨後一邊合成、一邊穿越 Sec61 進入 ER 腔,這就是共轉譯轉位。
SRP 的辨識本質上是一個結合平衡。若以 $K_d$ 描述 SRP 與訊號胜肽的解離常數,結合佔有率可寫成:
$$\theta = \frac{[\text{SRP}]}{[\text{SRP}] + K_d}$$
疏水性越強的訊號胜肽 $K_d$ 越小,$\theta$ 越接近 1,轉位效率越高。這解釋了為何訊號胜肽的疏水核心(h-region)在演化上高度保守——它直接決定第一道閘門的通過率。SRP 與其受體的交接還涉及兩者 GTPase 結構域的互鎌活化,GTP 水解提供方向性,避免空轉。
ER 腔內的折疊與品質管制:以動力學換取保真度
進入 ER 腔後,蛋白質面臨折疊、雙硫鍵形成與 N-連結醣基化。鈣聯蛋白/鈣網蛋白(calnexin/calreticulin)循環是一套以醣鏈為「成熟度標記」的監控系統:醣苷酶切除葡萄糖殘基,伴護蛋白判讀剩餘殘基決定保留或放行。折疊失敗者經 ERAD(ER-associated degradation) 逆轉位至胞質,由蛋白酶體降解。
品質管制的精髓在於「動力學校對(kinetic proofreading)」。設正確折疊蛋白離開 ER 的速率為 $k_{\text{exit}}$,錯誤蛋白被攔截降解的速率為 $k_{\text{deg}}$,則錯誤蛋白逃逸的機率約為:
$$P_{\text{escape}} \approx \frac{k_{\text{exit}}}{k_{\text{exit}} + k_{\text{deg}}}$$
細胞透過提高 $k_{\text{deg}}$(多次重複的伴護蛋白循環)把 $P_{\text{escape}}$ 壓低數個數量級,代價是時間與 ATP/GTP 消耗。這正是生物學常見的取捨:以能量與速度換取保真度。當未折疊蛋白堆積超過負荷,未折疊蛋白反應(UPR) 經 IRE1、PERK、ATF6 三條感測軸啟動,調降整體轉譯並上調伴護蛋白基因。
囊泡運輸:COPII、COPI 與膜的方向性流動
蛋白質離開 ER 需在 ER exit sites 被 COPII 外鞘捕捉成囊泡,向 Golgi 順行(anterograde)運輸;回收則由 COPI 負責逆行(retrograde)。COPII 的組裝由小 GTPase Sar1 啟動:GEF(Sec12)催化 Sar1 結合 GTP,使其兩親性 α 螺旋插入膜並募集外鞘蛋白,驅動膜彎曲成芽。
囊泡與目標膜的融合由 SNARE 蛋白配對完成,v-SNARE 與 t-SNARE 纏繞成四股螺旋束,把兩層膜拉近至可融合距離。融合的特異性與張力可視為一個自由能問題:SNARE 纏繞釋放的自由能 $\Delta G_{\text{SNARE}}$ 必須足以克服兩層膜近接的靜電與水合斥力能障 $\Delta G^{\ddagger}$。融合事件的速率近似遵循 Arrhenius 形式:
$$k_{\text{fusion}} = A \, e^{-\Delta G^{\ddagger}/RT}$$
多個 SNARE 複合體並聯可疊加自由能、降低有效能障,這也是融合「協同性」的物理根源。
定量小範例:分泌速率的限速步驟
假設某分泌蛋白在 ER 的折疊速率常數 $k_1 = 0.05\ \text{s}^{-1}$,COPII 包裝離開 ER 的速率常數 $k_2 = 0.20\ \text{s}^{-1}$。若視為兩步串聯一階反應,整體達到穩態前的「平均滯留時間」為各步驟時間常數之和:
$$\tau_{\text{total}} = \frac{1}{k_1} + \frac{1}{k_2} = \frac{1}{0.05} + \frac{1}{0.20} = 20 + 5 = 25\ \text{s}$$
可見折疊步驟(20 s)佔總滯留時間的 80%,是限速步驟(rate-limiting step)。這與實驗觀察一致:多數分泌蛋白的瓶頸在 ER 折疊與品質管制,而非囊泡運輸本身。若以伴護蛋白協助把 $k_1$ 提升至 $0.10\ \text{s}^{-1}$,則 $\tau_{\text{total}}$ 降為 $10 + 5 = 15\ \text{s}$,整體吞吐量提升約 40%——這正是細胞在高分泌需求(如漿細胞分泌抗體)時大幅擴張 ER 與伴護蛋白的理由。
Golgi 的分選與後續分泌
蛋白質穿越 Golgi 的順/反面堆疊(cis→trans)時,醣鏈被逐站修飾,形成如同「郵遞區號」的醣型。在 trans-Golgi network(TGN),蛋白質依標記分選進入不同囊泡:溶體酵素帶 甘露糖-6-磷酸(M6P) 標記被對應受體捕捉送往溶體;組成型分泌(constitutive)持續送往細胞膜;調節型分泌(regulated,如神經傳導物、激素)則濃縮儲存於分泌顆粒,待鈣訊號觸發胞吐。整個內膜系統因此是一個膜面積守恆的循環——順行運出的膜必須由逆行回收平衡,否則 ER 與 Golgi 會在數分鐘內被掏空或脹大。
深入探討(研究所視角)
主體把分泌途徑視為一條「串聯反應器」,但近年結構生物學與系統生物學揭示了更豐富的圖像。
轉位子的結構動態。 冷凍電顯(cryo-EM)已解析出 Sec61 與核糖體、TRAP、OST(oligosaccharyltransferase)複合體共組的「轉位子全酶」結構。Sec61 並非靜態通道,而是具有側向開口(lateral gate)的動態閘門:水溶性節段穿越中央孔徑進入 ER 腔,而疏水跨膜段則經由側向開口分配進脂雙層。這種「兩種出口」機制讓單一通道同時處理可溶分泌蛋白與多次跨膜蛋白的拓樸定向。近期研究進一步發現 EMC(ER membrane protein complex) 與 GET pathway 作為 Sec61 之外的平行插入途徑,專責尾錨定(tail-anchored)與棘手的跨膜蛋白,補足了「一個 translocon 走天下」舊模型的盲點。
膜接觸點與非囊泡運輸。 傳統模型強調囊泡,但 ER 與粒線體、質膜、Golgi 之間存在膜接觸點(membrane contact sites, MCS),由 tether 蛋白拉近至 10–30 nm,讓脂質與鈣離子可不經囊泡直接交換。VAP–OSBP 等脂質轉運蛋白在此以「逆向梯度」運送固醇,以磷酸肌醇(PI4P)的水解作為驅動的能量貨幣。這意味分泌途徑與脂質恆定是耦合的,不能孤立看待蛋白流。
單細胞與體學的異質性。 單細胞 RNA 定序與蛋白體學揭示,即使同型細胞,其分泌途徑容量(ER 大小、伴護蛋白表現、UPR 基調)也呈現顯著異質性。漿細胞分化過程中,XBP1 剪接驅動 ER 大規模擴張以支撐每秒數千分子的抗體輸出,是研究 UPR 與分泌容量耦合的經典模型。空間轉錄體學則開始把這些分泌表型對應到組織微環境。
系統生物學模型。 將分泌途徑建模為帶有品質管制回饋的反應–運輸網絡(reaction–transport network),可用常微分方程描述各胞器中折疊態、未折疊態與降解態的通量平衡,並以 metabolic control analysis 量化各步驟的「控制係數」。這類模型已被用於預測重組蛋白生產(如 CHO 細胞製藥)的瓶頸,指引以伴護蛋白或 UPR 調控因子進行的細胞工程。換言之,本文開頭那條多肽鏈的旅程,如今已能被寫成可模擬、可優化的方程組——這是分子細胞生物學邁向定量科學的縮影。