
膜電位與電化學梯度:從能斯特方程式到細胞的隱形電池
用電化學勢、能斯特與 GHK 方程式量化膜兩側的電壓,並延伸到結構、生物電與計算模型
從濃度差到電壓:膜兩側的隱形電池
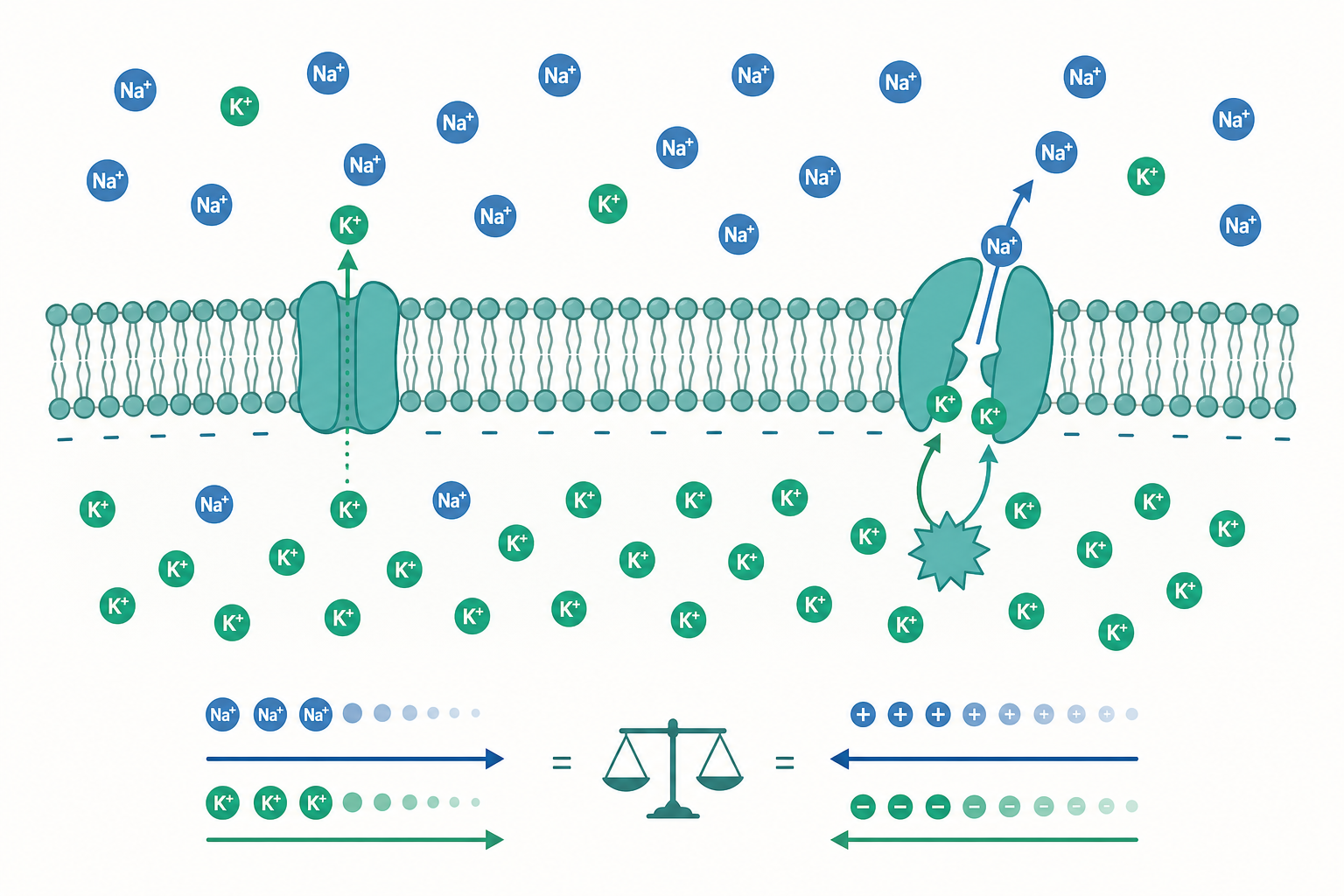
把一片活細胞的質膜想成一座極薄的水壩:兩側溶液看似平靜,實際上卻蓄積著巨大的勢能。任何時刻,靜止的神經元、肌肉細胞、甚至植物的保衛細胞,其膜兩側都維持著數十毫伏的電位差——這就是膜電位(membrane potential, $V_m$)。它並非生命的裝飾,而是訊號傳遞、能量轉換與物質運輸的共同貨幣。入門課程往往把它簡化成「鉀離子外流造成負電位」,但這句話既不夠精確,也藏著一整套電化學的量化邏輯。本篇要做的,是把這層直覺拆解成可計算的物理化學模型。
膜電位的根源是離子的不對稱分布加上膜的選擇性通透。細胞透過 Na⁺/K⁺-ATPase(鈉鉀幫浦)持續地把 3 個 Na⁺ 打出、2 個 K⁺ 拉進,建立起跨膜的離子濃度梯度。但濃度梯度本身不會自動變成電壓——還需要膜上對某種離子具選擇性的通道(如靜止時開放的 K⁺ 漏通道)。當 K⁺ 順著濃度梯度外流時,它帶走正電荷,使膜內側相對變負;這個負電位反過來吸引 K⁺ 留下。當「化學驅動力」(濃度差)與「電驅動力」(電位差)恰好抵消,淨流量為零,系統達到電化學平衡。
能斯特方程式:單一離子的平衡電位
對單一可滲透離子,達到平衡時的膜電位稱為該離子的平衡電位(equilibrium potential)或能斯特電位。其推導出發點是電化學勢(electrochemical potential)的相等。某離子 $X$ 在膜兩側的電化學勢分別為:
$$\tilde{\mu}_X = \mu_X^\circ + RT\ln[X] + zF\psi$$
其中 $R$ 為氣體常數($8.314\ \mathrm{J\,mol^{-1}K^{-1}}$)、$T$ 為絕對溫度、$z$ 為離子電荷數、$F$ 為法拉第常數($96485\ \mathrm{C\,mol^{-1}}$)、$\psi$ 為電位。令膜內外電化學勢相等並整理,即得能斯特方程式:
$$E_X = \frac{RT}{zF}\ln\frac{[X]_{out}}{[X]_{in}}$$
在生理溫度 $37^\circ\mathrm{C}$(310 K)下,把自然對數換成常用對數,可得便於記憶的形式:
$$E_X = \frac{61.5}{z}\log_{10}\frac{[X]_{out}}{[X]_{in}}\ \ (\mathrm{mV})$$
這條方程式的物理意義是:它給出「為了平衡某離子的濃度梯度,膜電位需要達到多少」。注意 $E_X$ 與離子的絕對濃度無關,只取決於比值——這是對數項的直接後果。
定量小範例:哺乳類神經元的 K⁺ 與 Na⁺ 平衡電位
以典型哺乳類神經元的離子濃度估算(單位 mM):
| 離子 | 細胞外 $[X]_{out}$ | 細胞內 $[X]_{in}$ |
|---|---|---|
| K⁺ | 5 | 140 |
| Na⁺ | 145 | 15 |
計算 $E_K$($z=+1$):
$$E_K = 61.5 \times \log_{10}\frac{5}{140} = 61.5 \times \log_{10}(0.0357)$$
$\log_{10}(0.0357) \approx -1.447$,故
$$E_K \approx 61.5 \times (-1.447) \approx -89\ \mathrm{mV}$$
計算 $E_{Na}$($z=+1$):
$$E_{Na} = 61.5 \times \log_{10}\frac{145}{15} = 61.5 \times \log_{10}(9.67) \approx 61.5 \times 0.985 \approx +61\ \mathrm{mV}$$
實測的靜止膜電位約為 $-70\ \mathrm{mV}$,落在 $E_K$($-89$)與 $E_{Na}$($+61$)之間,但明顯偏向 $E_K$。這提示我們:靜止膜對 K⁺ 通透性遠高於 Na⁺,但並非完全只透 K⁺。要描述「多種離子同時參與」的真實狀況,能斯特方程式就不夠用了。
GHK 方程式:多離子的加權平衡
當膜同時對多種離子通透,靜止膜電位由 Goldman–Hodgkin–Katz(GHK)方程式給出,它以各離子的相對通透係數 $P$ 為權重:
$$V_m = \frac{RT}{F}\ln\frac{P_K[K^+]_{out} + P_{Na}[Na^+]_{out} + P_{Cl}[Cl^-]_{in}}{P_K[K^+]_{in} + P_{Na}[Na^+]_{in} + P_{Cl}[Cl^-]_{out}}$$
注意 Cl⁻($z=-1$)的內外項位置與陽離子相反。當 $P_{Na}, P_{Cl} \to 0$,GHK 方程式退化回 K⁺ 的能斯特方程式——能斯特是 GHK 的特例。代入典型靜止態的通透比 $P_K : P_{Na} : P_{Cl} \approx 1 : 0.04 : 0.45$,可解出接近 $-70\ \mathrm{mV}$ 的結果。動作電位的本質,正是電壓門控 Na⁺ 通道瞬間開啟、使 $P_{Na}$ 暴增數百倍,膜電位被拉向 $E_{Na}$ 而去極化。
電化學梯度與驅動力:$\Delta G$ 的觀點
離子跨膜運輸的方向與能量,由電化學梯度決定。把一莫耳離子 $X$ 從膜外移到膜內的自由能變化為:
$$\Delta G = RT\ln\frac{[X]_{in}}{[X]_{out}} + zF V_m$$
第一項是濃度(化學)成分,第二項是電位(電學)成分。當 $\Delta G < 0$,離子自發內流;$\Delta G > 0$ 則需耗能主動運輸。對某離子而言,驅動力可簡潔表示為 $V_m - E_X$:當膜電位偏離該離子的平衡電位越遠,推動它流動的力量越大。例如靜止態 $V_m = -70$、$E_{Na} = +61$,Na⁺ 的驅動力高達 $-131\ \mathrm{mV}$(強烈內流),這也是 Na⁺ 通道一開、離子便奔流而入的原因。
這個 $\Delta G$ 框架還解釋了次級主動運輸:腸道上皮的 SGLT1 共運輸蛋白利用 Na⁺ 內流釋放的自由能,把葡萄糖逆濃度梯度拉進細胞——一個離子的電化學勢驅動另一分子的逆向運輸,能量帳本必須整體為負。粒線體內膜的質子動力勢(proton-motive force, pmf)同理:$\Delta p = \Delta\psi - \frac{2.3RT}{F}\Delta\mathrm{pH}$,把電位差與 pH 差合併成 ATP 合成酶的能量來源。膜電位,在這層意義上,是細胞通用的能量中介。
深入探討(研究所視角)
進入研究所層次,膜電位不再只是一個由 GHK 方程式描述的穩態純量,而是與離子通道的結構動力學、細胞的代謝體狀態及時空動態深度耦合的系統變數。
結構生物學的離子選擇性根源。 能斯特方程式假設膜對特定離子「選擇性通透」,但這選擇性的物理基礎直到 MacKinnon 團隊解出 KcsA 鉀通道晶體結構(1998,後獲 2003 年諾貝爾化學獎)才被原子級揭示。選擇性過濾器中由主鏈羰基氧(backbone carbonyl oxygen)形成的配位環境,精確模擬了水合 K⁺ 的氧殼層,使去水合的 K⁺ 通過的能障遠低於體積較小的 Na⁺——這是「越大的離子反而更易通過」這個反直覺現象的結構解。後續冷凍電鏡(cryo-EM)對電壓門控 Na⁺、Ca²⁺ 通道及其電壓感測域(voltage-sensing domain, S4 螺旋上的正電荷殘基)的高解析結構,使我們能把 GHK 中抽象的「通透係數 $P$」對應到具體的閘門構象轉換與門控電荷位移(gating charge)。
超越 GHK 的非平衡與電擴散模型。 GHK 方程式建立在「常電場假設」(constant field assumption)與穩態恆流之上,對狹窄孔道內的多離子佔據(multi-ion occupancy)、離子間排斥與能障跳躍並不準確。更嚴謹的處理採用 Poisson–Nernst–Planck(PNP)方程組,把離子通量寫成濃度梯度與電位梯度的耦合電擴散:$J = -D\left(\nabla c + \frac{zF}{RT}c\nabla\psi\right)$,並與描述電位分布的 Poisson 方程自洽求解。對於選擇性過濾器這類去水合、強相關的狹窄區域,PNP 的平均場近似仍不足,需引入分子動力學(MD)模擬或速率理論的能障模型。
單細胞與系統層次的膜電位體學。 近年電壓敏感螢光蛋白(genetically encoded voltage indicators, GEVIs,如 ASAP、Archon 系列)與全光學電生理(all-optical electrophysiology)讓研究者能在毫秒尺度同時記錄上千個神經元的膜電位動態,把單細胞電生理推向「電壓體學」的規模。與此並行,膜電位被重新認識為非神經細胞的訊號載體:Levin 等人的生物電(bioelectricity)研究指出,胚胎發育、再生與腫瘤生成過程中,組織層級的穩態膜電位模式($V_{mem}$ pattern)可作為形態發生的指令層,調控基因表現與細胞命運。這把原本侷限於可興奮細胞的膜電位概念,擴展為跨尺度的發育與病理變數。
代謝耦合與計算建模。 維持離子梯度的 Na⁺/K⁺-ATPase 消耗了神經元相當比例的 ATP,使膜電位與細胞能量狀態直接掛鉤;缺血或粒線體功能障礙導致幫浦失效時,梯度崩潰、麩胺酸過量釋放與興奮毒性隨之而來。在計算層面,Hodgkin–Huxley 模型(1952)以電壓門控電導的微分方程組首次定量重現了動作電位,至今仍是計算神經科學的基石;現代多腔室模型(multi-compartment models)則把 GHK、HH 動力學與離子幫浦、緩衝、擴散整合進空間離散的細胞幾何中,使膜電位研究從單點電壓邁向細胞與網路尺度的時空動態模擬。