
為什麼同一株菌,在燒瓶裡長得好,放大到五萬公升卻死一片?
從 Monod 動力學、chemostat 質量平衡到氧傳遞瓶頸,看生化工程如何把守恆定律套用在「活的反應器」上,並用優化學求解發酵放大的折衷。
為什麼同一株菌,在燒瓶裡長得好,放大到五萬公升卻死一片?
許多人第一次接觸生化工程(Biochemical Engineering),是從一個令人挫折的觀察開始:實驗室裡用 250 毫升搖瓶(shake flask)養大腸桿菌或酵母,菌長得又快又漂亮;可是一旦把同樣的菌種、同樣的培養基放進工廠的 50,000 公升發酵槽(fermenter),產率(yield)卻可能掉到剩下一半,甚至整批失敗。
問題不在生物學,而在工程。搖瓶裡的微生物沐浴在充足的氧氣與均勻的養分中;而在巨大的發酵槽裡,氧氣要從氣泡溶進液體、再擴散到每一個細胞,這條路徑變得非常漫長。同時,攪拌槳產生的剪切力(shear stress)、局部養分濃度不均、代謝產生的熱量無法即時帶走——這些「輸送現象」(transport phenomena)才是放大(scale-up)成敗的關鍵。
生化工程,正是把化學工程的質量守恆、動量守恆與能量守恆,套用在「活的反應器」上。微生物不是被動的觸媒,牠們會生長、會死亡、會改變代謝路徑——這讓生化反應器(bioreactor)的設計,比一般化學反應器多了一層動態的生命邏輯。
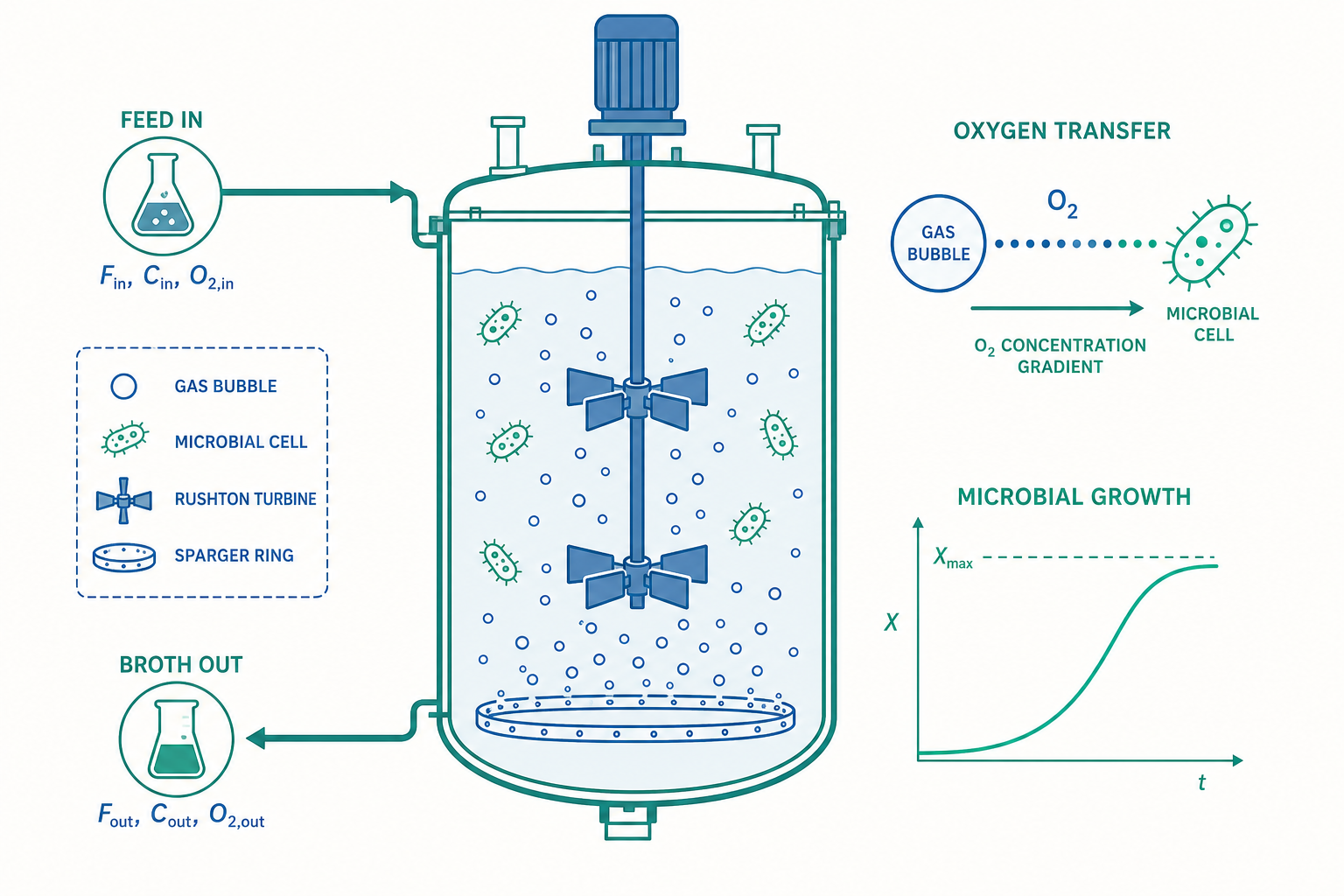
細胞生長動力學:把「活的觸媒」寫成方程式
要設計發酵程序,第一步是用數學描述細胞如何生長。最經典的是 Monod 方程式,它把比生長速率(specific growth rate)$\mu$ 與限制性基質(limiting substrate)濃度 $S$ 連結起來:
$$\mu = \mu_{\max} \frac{S}{K_S + S}$$
其中 $\mu_{\max}$ 是最大比生長速率(單位 $\text{h}^{-1}$),$K_S$ 是半飽和常數(saturation constant),代表 $\mu = \tfrac{1}{2}\mu_{\max}$ 時的基質濃度。這條曲線的形狀和酵素動力學的 Michaelis–Menten 方程式如出一轍——這並非巧合,因為細胞生長的速率最終受限於內部酵素反應。
在批次培養(batch culture)中,細胞濃度 $X$ 隨時間呈指數增長:
$$\frac{dX}{dt} = \mu X \quad\Longrightarrow\quad X(t) = X_0 \, e^{\mu t}$$
當基質充足($S \gg K_S$)時,$\mu \approx \mu_{\max}$,細胞進入對數生長期(exponential phase)。隨著基質耗盡,$\mu$ 下降,最終進入靜止期(stationary phase)。
我們還需要兩個重要的得率係數(yield coefficient),把不同物質的消耗與生成綁在一起:
$$Y_{X/S} = -\frac{dX}{dS}, \qquad Y_{P/S} = -\frac{dP}{dS}$$
$Y_{X/S}$ 是「每消耗 1 克基質能產生多少克細胞」,$Y_{P/S}$ 則是基質轉化為目標產物 $P$ 的效率。這些係數讓我們能在質量平衡中把基質、菌體、產物三者串起來。
質量平衡:發酵槽的會計帳
生化工程的核心工具,是對反應器內每一種物質做質量平衡。以連續攪拌槽反應器(CSTR)型態運轉的連續培養(continuous culture, chemostat) 為例,假設體積 $V$ 固定、進料流量 $F$ 固定,定義稀釋率(dilution rate)$D = F/V$(單位 $\text{h}^{-1}$)。
對細胞濃度 $X$ 做平衡,進料無菌($X_{in}=0$):
$$\underbrace{V\frac{dX}{dt}}_{\text{累積}} = \underbrace{F \cdot 0}_{\text{流入}} - \underbrace{F X}_{\text{流出}} + \underbrace{\mu X V}_{\text{生成}}$$
兩邊除以 $V$:
$$\frac{dX}{dt} = \mu X - D X$$
在穩態(steady state,$dX/dt = 0$)且 $X \neq 0$ 時,得到一個極簡潔而深刻的結論:
$$\boxed{\mu = D}$$
也就是說,在 chemostat 中,操作者只要設定進料流速,就等於設定了細胞的生長速率。這是生化工程最優雅的結果之一:我們用一個外部可控的工程參數($D$),鎖定了生物體內在的生理狀態($\mu$)。
對基質 $S$ 同樣做平衡(進料濃度 $S_0$,消耗速率為 $\mu X / Y_{X/S}$):
$$\frac{dS}{dt} = D(S_0 - S) - \frac{\mu X}{Y_{X/S}}$$
穩態時聯立 $\mu = D$ 與 Monod 方程式,可解出穩態基質濃度:
$$S = \frac{K_S \, D}{\mu_{\max} - D}$$
並進一步得到穩態菌體濃度:
$$X = Y_{X/S}\,(S_0 - S) = Y_{X/S}\left(S_0 - \frac{K_S D}{\mu_{\max} - D}\right)$$
注意:當 $D$ 逼近 $\mu_{\max}$ 時,$S$ 急遽上升、$X$ 趨近於零——此時細胞來不及生長就被沖出反應器,稱為沖洗(washout)。臨界稀釋率為:
$$D_{\text{crit}} = \mu_{\max}\frac{S_0}{K_S + S_0}$$
設計連續發酵時,工程師必須讓 $D$ 安全地低於 $D_{\text{crit}}$。
氧氣輸送:放大失敗的頭號元兇
回到開頭的謎題。好氧(aerobic)發酵中,氧氣的供應幾乎總是放大的瓶頸,原因是氧在水中的溶解度極低(25°C、1 atm 空氣下飽和溶氧僅約 7–8 mg/L)。氧氣的供應速率以氧氣傳遞速率(Oxygen Transfer Rate, OTR)描述:
$$\text{OTR} = k_L a \,(C^* - C_L)$$
其中 $k_L a$ 是體積氧傳遞係數(volumetric mass transfer coefficient,單位 $\text{h}^{-1}$),$C^*$ 是與氣相平衡的飽和溶氧濃度,$C_L$ 是液相實際溶氧濃度。$k_L$ 是液膜傳質係數,$a$ 是單位體積的氣液界面積——攪拌得越劇烈、氣泡越小越多,$a$ 越大,$k_L a$ 越高。
而細胞的耗氧速率(Oxygen Uptake Rate, OUR)為:
$$\text{OUR} = q_{O_2} X$$
$q_{O_2}$ 是比耗氧速率。發酵能否維持的鐵律是:
$$\text{OTR} \geq \text{OUR} \quad\Longrightarrow\quad k_L a\,(C^* - C_L) \geq q_{O_2} X$$
一旦菌體濃度 $X$ 太高、$k_L a$ 跟不上,$C_L$ 就會跌到臨界溶氧(critical dissolved oxygen)以下,細胞缺氧、代謝轉向(例如酵母進行 Crabtree 效應產生乙醇),產率崩潰。
這正是為什麼放大如此困難:搖瓶靠表面氣液交換,$k_L a$ 自然夠用;但發酵槽體積放大後,表面積與體積比急遽下降,必須靠強力攪拌與通氣(sparging)來補足 $k_L a$。然而攪拌功率有上限、剪切力會傷害細胞——這是一個典型的工程取捨。
攪拌功率與放大準則:到底要固定哪個量?
通氣攪拌槽的攪拌功率,未通氣時可用功率準數(power number)$N_p$ 估算:
$$P = N_p \, \rho \, N^3 D_i^5$$
其中 $\rho$ 為液體密度、$N$ 為攪拌轉速(rev/s)、$D_i$ 為攪拌槳直徑。放大時,工程師面臨一個根本難題:無法同時保持所有無因次群相同。常見的放大準則包括:
- 固定單位體積功率 $P/V$(最常用於好氧發酵,因為它與 $k_L a$ 高度相關)
- 固定 $k_L a$(直接守住氧傳遞)
- 固定槳尖速度 $\pi N D_i$(控制剪切力,保護敏感的動物細胞或菌絲體)
- 固定混合時間(mixing time)
這幾個準則彼此衝突:若放大時固定 $P/V$,則槳尖速度會上升、剪切力增加;若固定槳尖速度,則 $P/V$ 會下降、氧傳遞變差。沒有完美解,只有針對特定生物系統的最佳折衷——這就是把優化學(optimization)導入生化工程的切入點:以氧傳遞約束、剪切容忍度、能耗成本為限制式,求解最適的攪拌與通氣組合。
看一個例子
某好氧發酵欲將大腸桿菌培養到菌體濃度 $X = 20\ \text{g/L}$。已知比耗氧速率 $q_{O_2} = 0.4\ \text{g O}_2 / (\text{g cell} \cdot \text{h})$,飽和溶氧 $C^* = 7.5\ \text{mg/L}$,欲將液相溶氧維持在 $C_L = 2.0\ \text{mg/L}$(即臨界值之上)。試問所需的最低 $k_L a$ 為何?
第一步:計算耗氧速率 OUR。
$$\text{OUR} = q_{O_2} X = 0.4 \times 20 = 8.0\ \text{g O}_2/(\text{L}\cdot\text{h}) = 8000\ \text{mg/(L}\cdot\text{h})$$
第二步:由 OTR ≥ OUR 反推 $k_L a$。 為維持穩態,令 $\text{OTR} = \text{OUR}$:
$$k_L a = \frac{\text{OUR}}{C^* - C_L} = \frac{8000}{7.5 - 2.0} = \frac{8000}{5.5} \approx 1455\ \text{h}^{-1}$$
第三步:工程判讀。 一般工業攪拌發酵槽的 $k_L a$ 約落在 $100\sim 500\ \text{h}^{-1}$。我們算出需要約 $1455\ \text{h}^{-1}$,遠超常規上限——這代表單靠普通攪拌通氣無法在此菌體濃度維持供氧。可行對策包括:採用富氧空氣(提高 $C^*$)、提高槽壓(亨利定律使 $C^*$ 上升)、改用分批饋料(fed-batch)控制 $X$ 的上升速度,或降低目標菌體濃度。
這個例子說明:生化工程的設計不是「把菌養越多越好」,而是在氧傳遞的物理極限內,找到產率與可操作性的平衡點。
重點回顧
- 生化工程=把質能守恆套用在活的反應器上。 微生物會生長、死亡、改變代謝,使生化反應器比化學反應器多了動態的生命邏輯,放大失敗多半源於輸送現象而非生物學本身。
- Monod 方程式描述生長動力學,$\mu = \mu_{\max} S/(K_S+S)$,形式上與 Michaelis–Menten 一致;得率係數 $Y_{X/S}$、$Y_{P/S}$ 把基質、菌體、產物串入質量平衡。
- 連續培養(chemostat)的黃金結論是 $\mu = D$:操作者用稀釋率直接鎖定細胞生長速率,但 $D$ 必須低於臨界稀釋率以避免沖洗(washout)。
- 氧傳遞是好氧放大的瓶頸:必須滿足 $k_L a (C^*-C_L) \geq q_{O_2} X$,搖瓶到大槽的成敗,關鍵在 $k_L a$ 能否跟上菌體濃度。
- 放大準則互相衝突(固定 $P/V$ 或槳尖速度或 $k_L a$ 不可兼得),最終必須借助優化學在氧傳遞、剪切、能耗等約束下求折衷。
深入探討(研究所視角)
研究所階段的生化工程,會在前述穩態框架上疊加幾層更貼近真實的複雜性,值得在此勾勒。
一、結構化與分群動力學模型(structured / segregated models)。 Monod 模型把細胞當成一個沒有內部結構的「黑箱」(unstructured),且假設群體均一(unsegregated)。但真實細胞具有內部狀態(DNA、RNA、酵素、儲存物質),且群體在年齡與大小上具分布。結構化模型(如 cybernetic models)追蹤胞內成分,能描述 diauxic growth(雙基質次序利用)等現象;族群平衡模型(population balance equations)則以分布函數描述群體異質性,需求解積分–偏微分方程。
二、CFD 耦合的非理想混合。 工業大槽絕非理想全混合,存在養分與溶氧的空間梯度。現代做法是將計算流體力學(CFD)求得的流場,與細胞動力學耦合,模擬細胞在槽內循環時反覆經歷高糖/低氧、低糖/高氧區域的「生命歷程」(lifeline analysis)。這解釋了放大時觀察到的產率損失,並指引攪拌槳配置的優化。
三、代謝通量分析與 fed-batch 最佳控制。 將胞內代謝網路寫成穩態化學計量矩陣 $\mathbf{S}$,在 $\mathbf{S}\,\mathbf{v}=0$ 的約束下,以線性規劃最大化目標通量,即通量平衡分析(Flux Balance Analysis, FBA)。在程序層面,分批饋料(fed-batch)的饋料曲線本身是一個動態最佳化問題:
$$\max_{F(t)} \; P(t_f) \quad \text{s.t.}\quad \frac{dX}{dt},\frac{dS}{dt},\frac{dP}{dt}=f(\cdot),\; \text{OTR}\geq\text{OUR},\; 0\leq F\leq F_{\max}$$
可用 Pontryagin 最大原理或數值最佳控制求解最適饋料策略,這正是優化學與生化工程交會的前沿。
四、下游分離(downstream processing)。 發酵只是上游;產物純化(細胞分離、破菌、層析、結晶)的成本常占整廠的 50–80%。下游與上游必須協同設計,例如選擇分泌型表達以簡化純化,這呼應了化學工程「反應與分離整合」的主題群精神。
從一只搖瓶到一座工廠,生化工程要回答的始終是同一個問題:如何在輸送現象的物理極限內,讓活的觸媒穩定、可控、可放大地工作。